

Pregnenolone sulphate-independent inhibition of TRPM3 channels by progesterone
- PMID: 22000496
- PMCID: PMC3275754
- DOI: 10.1016/j.ceca.2011.09.005
Pregnenolone sulphate-independent inhibition of TRPM3 channels by progesterone
Abstract
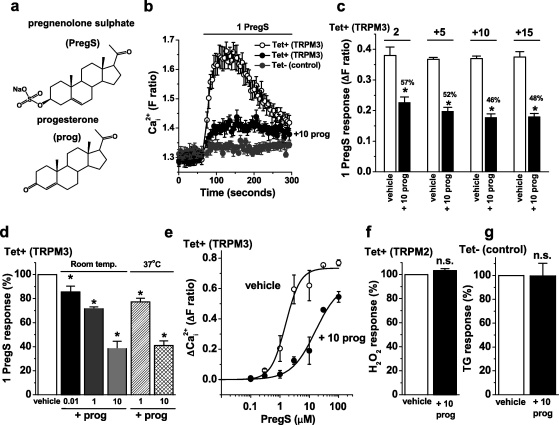
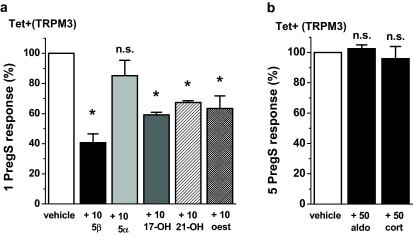
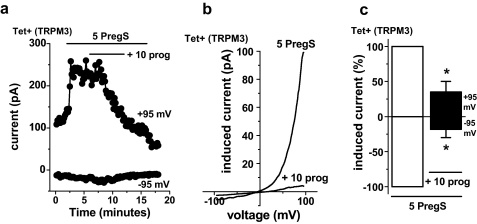
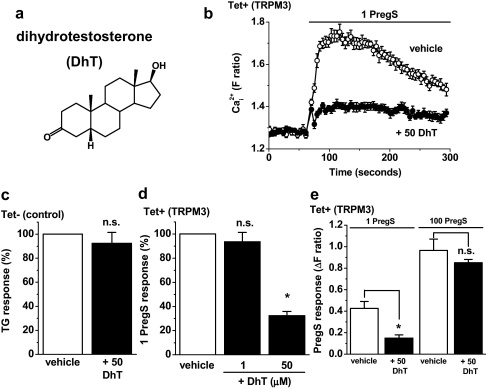
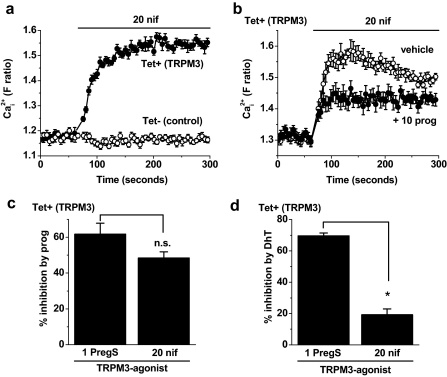
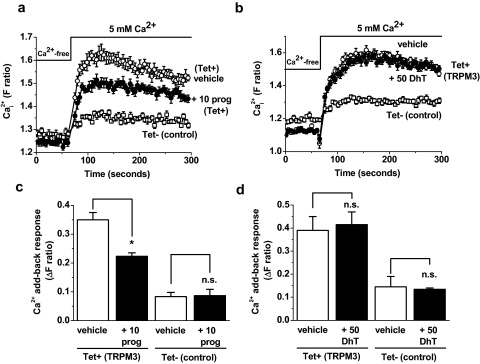
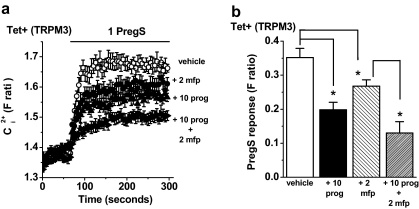
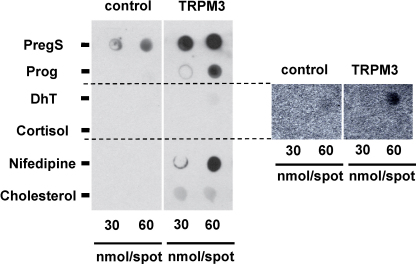
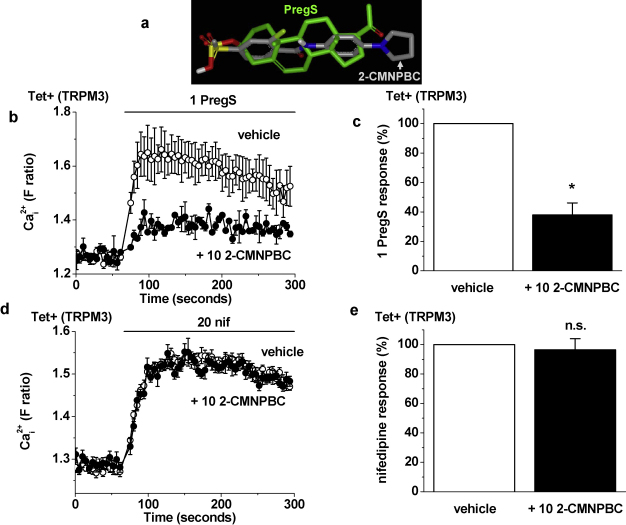
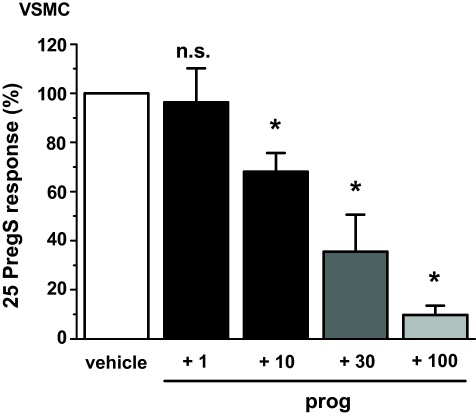
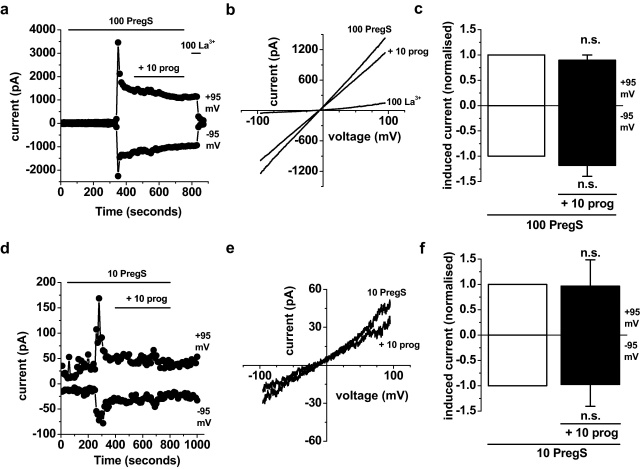
Transient Receptor Potential Melastatin 3 (TRPM3) is a widely expressed calcium-permeable non-selective cation channel that is stimulated by high concentrations of nifedipine or by physiological steroids that include pregnenolone sulphate. Here we sought to identify steroids that inhibit TRPM3. Channel activity was studied using calcium-measurement and patch-clamp techniques. Progesterone (0.01-10μM) suppressed TRPM3 activity evoked by pregnenolone sulphate. Progesterone metabolites and 17β-oestradiol were also inhibitory but the effects were relatively small. Dihydrotestosterone was an inhibitor at concentrations higher than 1μM. Corticosteroids lacked effect. Overlay assays indicated that pregnenolone sulphate, progesterone and dihydrotestosterone bound to TRPM3. In contrast to dihydrotestosterone, progesterone inhibited nifedipine-evoked TRPM3 activity or activity in the absence of an exogenous activator, suggesting a pregnenolone sulphate-independent mechanism of action. Dihydrotestosterone, like a non-steroid look-alike compound, acted as a competitive antagonist at the pregnenolone sulphate binding site. Progesterone inhibited endogenous TRPM3 in vascular smooth muscle cells. Relevance of TRPM3 or the progesterone effect to ovarian cells, which have been suggested to express TRPM3, was not identified. The data further define a chemical framework for competition with pregnenolone sulphate at TRPM3 and expand knowledge of steroid interactions with TRPM3, suggesting direct steroid binding and pregnenolone sulphate-independent inhibition by progesterone.
Crown Copyright © 2011. Published by Elsevier India Pvt Ltd. All rights reserved.
Figures
Similar articles
-
Sex difference in TRPM3 channel functioning in nociceptive and vascular systems: an emerging target for migraine therapy in females?J Headache Pain. 2025 Feb 24;26(1):40. doi: 10.1186/s10194-025-01966-9. J Headache Pain. 2025. PMID: 39994546 Free PMC article. Review.
-
Structural requirements of steroidal agonists of transient receptor potential melastatin 3 (TRPM3) cation channels.Br J Pharmacol. 2014 Feb;171(4):1019-32. doi: 10.1111/bph.12521. Br J Pharmacol. 2014. PMID: 24251620 Free PMC article.
-
Stereo-selective inhibition of transient receptor potential TRPC5 cation channels by neuroactive steroids.Br J Pharmacol. 2011 Apr;162(7):1509-20. doi: 10.1111/j.1476-5381.2010.01136.x. Br J Pharmacol. 2011. PMID: 21108630 Free PMC article.
-
Cis-isomerism and other chemical requirements of steroidal agonists and partial agonists acting at TRPM3 channels.Br J Pharmacol. 2010 Sep;161(2):430-41. doi: 10.1111/j.1476-5381.2010.00892.x. Br J Pharmacol. 2010. PMID: 20735426 Free PMC article.
-
Signal transduction via TRPM3 channels in pancreatic β-cells.J Mol Endocrinol. 2013 Apr 23;50(3):R75-83. doi: 10.1530/JME-12-0237. Print 2013 Jun. J Mol Endocrinol. 2013. PMID: 23511953 Review.
Cited by
-
Activation and inhibition of transient receptor potential TRPM3-induced gene transcription.Br J Pharmacol. 2014 May;171(10):2645-58. doi: 10.1111/bph.12524. Br J Pharmacol. 2014. PMID: 24895737 Free PMC article.
-
Diclofenac, a nonsteroidal anti-inflammatory drug, is an antagonist of human TRPM3 isoforms.Pharmacol Res Perspect. 2016 Apr 7;4(3):e00232. doi: 10.1002/prp2.232. eCollection 2016 Jun. Pharmacol Res Perspect. 2016. PMID: 27433342 Free PMC article.
-
Mechanosensitive receptors in migraine: a systematic review.J Headache Pain. 2024 Jan 15;25(1):6. doi: 10.1186/s10194-023-01710-1. J Headache Pain. 2024. PMID: 38221631 Free PMC article.
-
TRPM3 in Brain (Patho)Physiology.Front Cell Dev Biol. 2021 Feb 26;9:635659. doi: 10.3389/fcell.2021.635659. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33732703 Free PMC article. Review.
-
Sex difference in TRPM3 channel functioning in nociceptive and vascular systems: an emerging target for migraine therapy in females?J Headache Pain. 2025 Feb 24;26(1):40. doi: 10.1186/s10194-025-01966-9. J Headache Pain. 2025. PMID: 39994546 Free PMC article. Review.
References
-
- Wagner T.F., Loch S., Lambert S., Straub I., Mannebach S., Mathar I., Dufer M., Lis A., Flockerzi V., Philipp S.E., Oberwinkler J. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic beta cells. Nat. Cell Biol. 2008;10:1421–1430. - PubMed
-
- Hoffmann A., Grimm C., Kraft R., Goldbaum O., Wrede A., Nolte C., Hanisch U.K., Richter-Landsberg C., Bruck W., Kettenmann H., Harteneck C. TRPM3 is expressed in sphingosine-responsive myelinating oligodendrocytes. J. Neurochem. 2010;114:654–665. - PubMed
-
- Naylor J., Li J., Milligan C.J., Zeng F., Sukumar P., Hou B., Sedo A., Yuldasheva N., Majeed Y., Beri D., Jiang S., Seymour V.A., McKeown L., Kumar B., Harteneck C., O‘Regan D., Wheatcroft S.B., Kearney M.T., Jones C., Porter K.E., Beech D.J. Pregnenolone sulphate- and cholesterol-regulated TRPM3 channels coupled to vascular smooth muscle secretion and contraction. Circ. Res. 2010;106:1507–1515. - PMC - PubMed
-
- Vriens J., Owsianik G., Hofmann T., Philipp S.E., Stab J., Chen X., Benoit M., Xue F., Janssens A., Kerselaers S., Oberwinkler J., Vennekens R., Gudermann T., Nilius B., Voets T. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron. 2011;70:482–494. - PubMed
-
- Wagner T.F., Drews A., Loch S., Mohr F., Philipp S.E., Lambert S., Oberwinkler J. TRPM3 channels provide a regulated influx pathway for zinc in pancreatic beta cells. Pflugers Arch. 2010;460:755–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
