

Bidirectional resection of DNA double-strand breaks by Mre11 and Exo1
- PMID: 22002605
- PMCID: PMC3214165
- DOI: 10.1038/nature10515
Bidirectional resection of DNA double-strand breaks by Mre11 and Exo1
Abstract
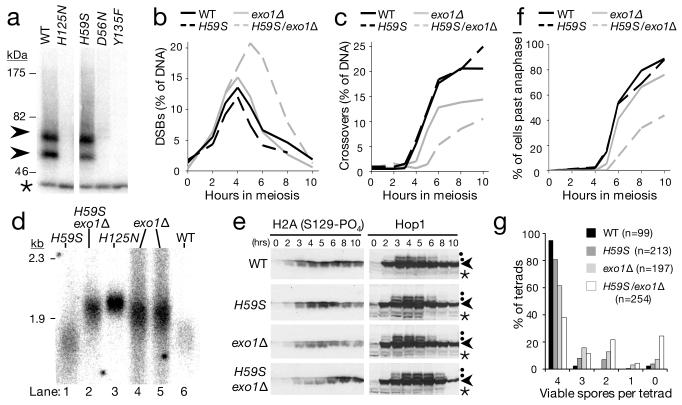
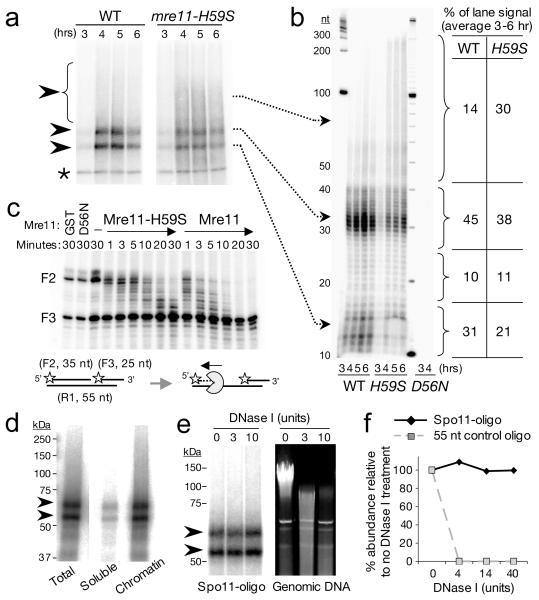
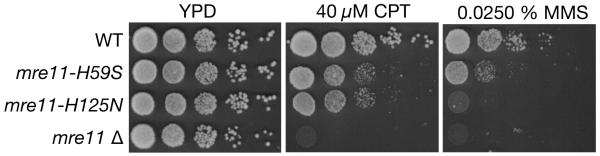
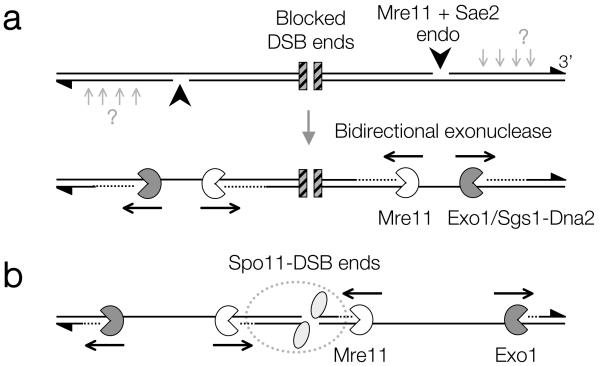
Repair of DNA double-strand breaks (DSBs) by homologous recombination requires resection of 5'-termini to generate 3'-single-strand DNA tails. Key components of this reaction are exonuclease 1 and the bifunctional endo/exonuclease, Mre11 (refs 2-4). Mre11 endonuclease activity is critical when DSB termini are blocked by bound protein--such as by the DNA end-joining complex, topoisomerases or the meiotic transesterase Spo11 (refs 7-13)--but a specific function for the Mre11 3'-5' exonuclease activity has remained elusive. Here we use Saccharomyces cerevisiae to reveal a role for the Mre11 exonuclease during the resection of Spo11-linked 5'-DNA termini in vivo. We show that the residual resection observed in Exo1-mutant cells is dependent on Mre11, and that both exonuclease activities are required for efficient DSB repair. Previous work has indicated that resection traverses unidirectionally. Using a combination of physical assays for 5'-end processing, our results indicate an alternative mechanism involving bidirectional resection. First, Mre11 nicks the strand to be resected up to 300 nucleotides from the 5'-terminus of the DSB--much further away than previously assumed. Second, this nick enables resection in a bidirectional manner, using Exo1 in the 5'-3' direction away from the DSB, and Mre11 in the 3'-5' direction towards the DSB end. Mre11 exonuclease activity also confers resistance to DNA damage in cycling cells, suggesting that Mre11-catalysed resection may be a general feature of various DNA repair pathways.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
