

Review
doi: 10.1104/pp.111.187088.
Epub 2011 Oct 14.
The regulatory activities of plant microRNAs: a more dynamic perspective
Affiliations
- PMID: 22003084
- PMCID: PMC3327222
- DOI: 10.1104/pp.111.187088
Item in Clipboard
Review
The regulatory activities of plant microRNAs: a more dynamic perspective
Plant Physiol.
2011 Dec.
No abstract available
Figures
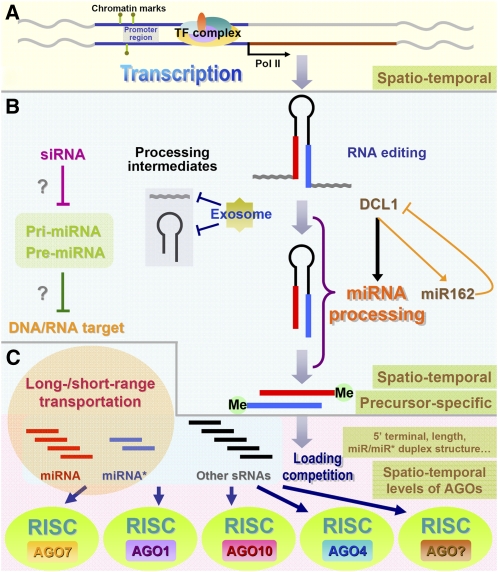
Schematic presentation showing the dynamic nature of miRNA biogenesis in plants. A, RNA polymerase II (Pol II)-dependent transcription of the miRNA genes. Chromatin marks including DNA methylation and histone modifications, and the combinatory regulation of many TFs, together contribute the spatiotemporal expression patterns of the miRNA genes in plants. B, Processing of miRNA precursors and miRNA maturation. RNA editing on the miRNA precursors plays a role in changing the original sequence information encoded by the miRNA gene loci. The exosome was suggested to be implicated in digesting the processing intermediates from the miRNA precursors, ensuring relatively high processing efficiency. The processing efficiency also shows a high precursor sequence-specific dependence. Methylation at the 3′ ends of the miRNA/miRNA* duplex (“Me” here represents the methyl group) is crucial for the stabilization of miRNA and miRNA*. A feedback circuit between DCL1 and miR162 exists within the processing procedure. Moreover, the siRNAs complementary to specific pri-miRNAs and pre-miRNAs exhibit a potential repressive role in miRNA processing. More interestingly, the pri-miRNAs and pre-miRNAs may possess their own targets. C, Sorting into the AGO-associated miRISCs. A drastic loading competition may exist among miRNAs, miRNA*s, and other sRNA species. Not all the miRNAs are incorporated into the AGO1 complex. The 5′ terminal composition and the sequence length of the miRNAs, the structure of the miRNA/miRNA* duplex, and other undetermined factors have a significant influence on the loading patterns of the mature miRNAs.
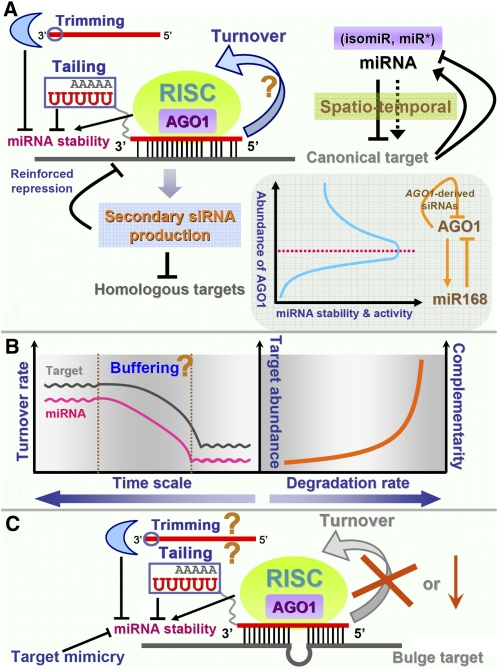
Schematic summarization of the factors influencing miRNA action and in vivo levels. A, Based on recent reports in animals, the turnover use of the mature miRNAs is proposed in plants, which needs further validation (denoted by the question mark). The 3′ tailing and the 3′-to-5′ trimming greatly affect the stability of mature miRNAs. Certain miRNA targets, such as TF genes, could in turn regulate the miRNAs, thus forming feedback regulatory circuits. The secondary siRNAs amplified from the cleaved target transcripts of a specific miRNA could reinforce miRNA-mediated gene silencing posttranscriptionally. One example is provided by the feedback circuit between AGO1 and miR168 in Arabidopsis. The secondary siRNAs derived from miR168-cleaved AGO1 transcripts could further regulate the expression of AGO1 posttranscriptionally. On the other hand, the abundance of the AGO1 protein significantly affects the activities and the stability of numerous miRNAs. B, The adjustable turnover rate of certain miRNAs may form an elaborate buffering system within the miRNA-mediated regulatory networks, which needs further investigation (denoted by the question mark in the left panel). Based on current hints in plants and animals, a target abundance- and complementarity-dependent model was proposed to be implicated in modulating the miRNA degradation rate (right panel). C, Based on the phenomenon of target mimicry observed by Franco-Zorrilla et al. (2007), the miRNA could be sequestered by a target decoy with a central bulge within the target recognition sites. Thus, miRNA turnover will be inhibited in that case. Although it is still not clear, the 3′ tailing and 3′-to-5′ trimming may also occur on the miRNAs sequestered by the bulged targets (denoted by question marks).
References
-
- Allen E, Howell MD. (2010) miRNAs in the biogenesis of trans-acting siRNAs in higher plants. Semin Cell Dev Biol 21: 798–804 - PubMed
-
- Allen E, Xie Z, Gustafson AM, Carrington JC. (2005) MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221 - PubMed
-
- Allen E, Xie Z, Gustafson AM, Sung GH, Spatafora JW, Carrington JC. (2004) Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat Genet 36: 1282–1290 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
