

PCNA binding domains in all three subunits of yeast DNA polymerase δ modulate its function in DNA replication
- PMID: 22003126
- PMCID: PMC3207673
- DOI: 10.1073/pnas.1109981108
PCNA binding domains in all three subunits of yeast DNA polymerase δ modulate its function in DNA replication
Abstract
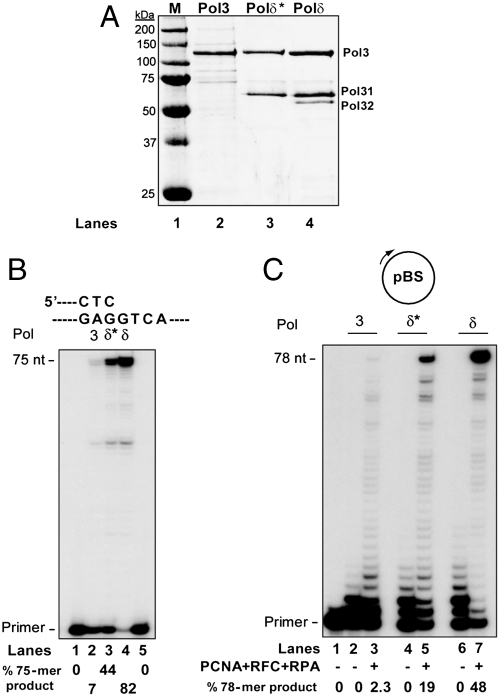
DNA polymerase δ (Polδ) plays an essential role in replication from yeast to humans. Polδ in Saccharomyces cerevisiae is comprised of three subunits, the catalytic subunit Pol3 and the accessory subunits Pol31 and Pol32. Yeast Polδ exhibits a very high processivity in synthesizing DNA with the proliferating cell nuclear antigen (PCNA) sliding clamp; however, it has remained unclear how Polδ binds PCNA to achieve its high processivity. Here we show that PCNA interacting protein (PIP) motifs in all three subunits contribute to PCNA-stimulated DNA synthesis by Polδ, and mutational inactivation of all three PIP motifs abrogates its ability to synthesize DNA with PCNA. Genetic analyses of mutations in these PIPs have revealed that in the absence of functional Pol32 PIP domain, PCNA binding by both the Pol3 and Pol31 subunits becomes essential for cell viability. Based on our biochemical and genetic studies we infer that yeast Polδ can simultaneously utilize all three PIP motifs during PCNA-dependent DNA synthesis, and suggest that Polδ binds the PCNA homotrimer via its three subunits. We consider the implications of these observations for Polδ's role in DNA replication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Waga S, Stillman B. The DNA replication fork in eukaryotic cells. Annu Rev Biochem. 1998;67:721–751. - PubMed
-
- Tsurimoto T, Stillman B. Replication factors required for SV40 DNA replication in vitro II. Switching of DNA polymerase alpha and delta during initiation of leading and lagging strand synthesis. J Biol Chem. 1991;266:1961–1968. - PubMed
-
- Waga S, Stillman B. Anatomy of a DNA replication fork revealed by reconstitution of SV40 DNA replication in vitro. Nature. 1994;369:207–212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
