

Two distinct catalytic strategies in the hepatitis δ virus ribozyme cleavage reaction
- PMID: 22003985
- PMCID: PMC3220057
- DOI: 10.1021/bi201157t
Two distinct catalytic strategies in the hepatitis δ virus ribozyme cleavage reaction
Abstract
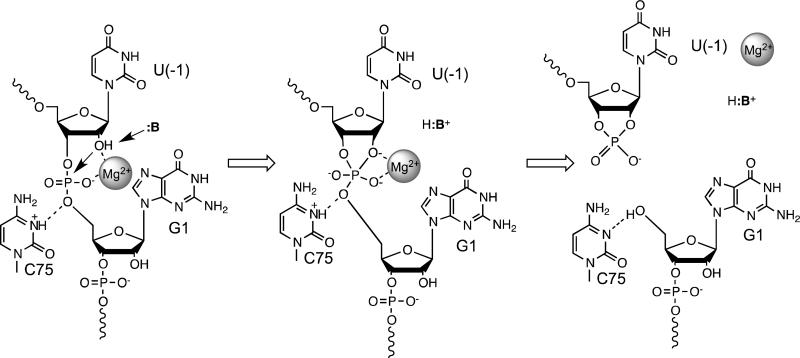
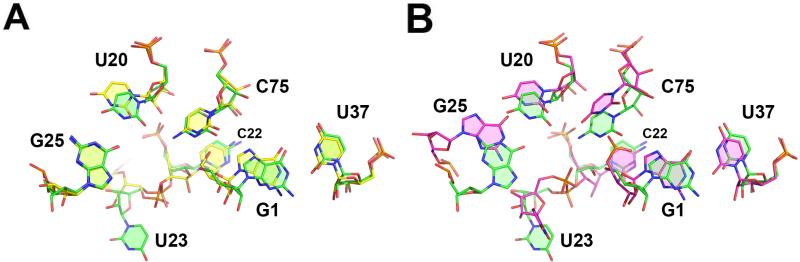
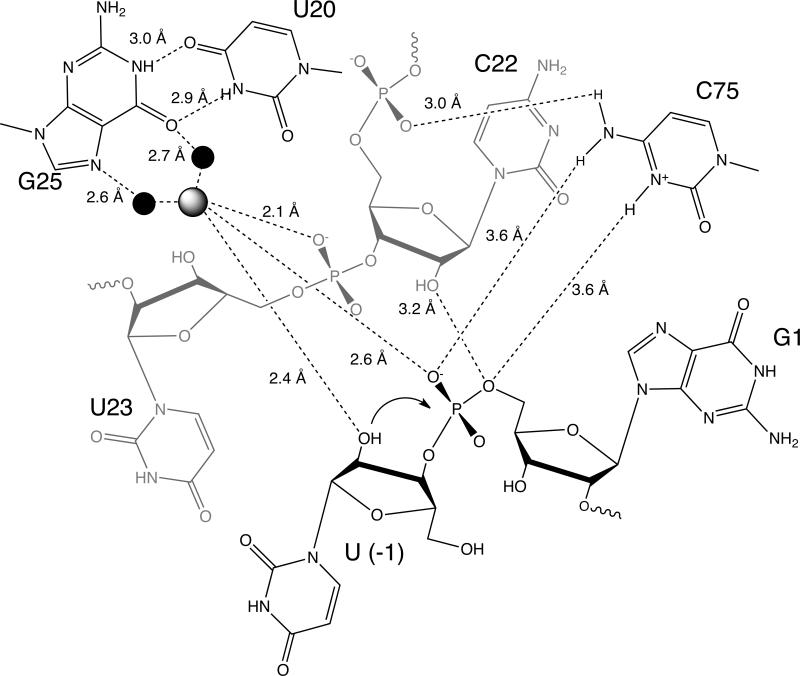
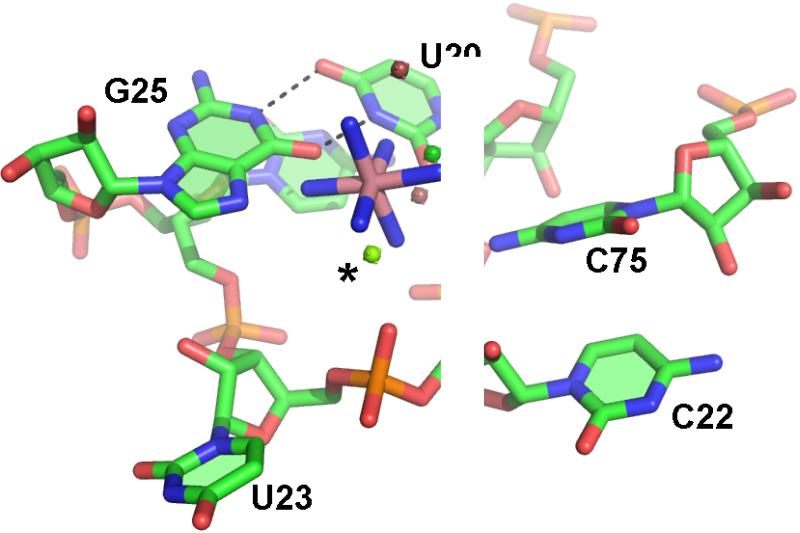
The hepatitis delta virus (HDV) ribozyme and related RNAs are widely dispersed in nature. This RNA is a small nucleolytic ribozyme that self-cleaves to generate products with a 2',3'-cyclic phosphate and a free 5'-hydroxyl. Although small ribozymes are dependent on divalent metal ions under biologically relevant buffer conditions, they function in the absence of divalent metal ions at high ionic strengths. This characteristic suggests that a functional group within the covalent structure of small ribozymes is facilitating catalysis. Structural and mechanistic analyses have demonstrated that the HDV ribozyme active site contains a cytosine with a perturbed pK(a) that serves as a general acid to protonate the leaving group. The reaction of the HDV ribozyme in monovalent cations alone never approaches the velocity of the Mg(2+)-dependent reaction, and there is significant biochemical evidence that a Mg(2+) ion participates directly in catalysis. A recent crystal structure of the HDV ribozyme revealed that there is a metal binding pocket in the HDV ribozyme active site. Modeling of the cleavage site into the structure suggested that this metal ion can interact directly with the scissile phosphate and the nucleophile. In this manner, the Mg(2+) ion can serve as a Lewis acid, facilitating deprotonation of the nucleophile and stabilizing the conformation of the cleavage site for in-line attack of the nucleophile at the scissile phosphate. This catalytic strategy had previously been observed only in much larger ribozymes. Thus, in contrast to most large and small ribozymes, the HDV ribozyme uses two distinct catalytic strategies in its cleavage reaction.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
