

Review
doi: 10.1021/cr200187e.
Epub 2011 Oct 17.
Uncovering ubiquitin and ubiquitin-like signaling networks
Affiliations
- PMID: 22004258
- PMCID: PMC3238414
- DOI: 10.1021/cr200187e
Item in Clipboard
Review
Uncovering ubiquitin and ubiquitin-like signaling networks
Chem Rev.
.
Free PMC article
No abstract available
Figures
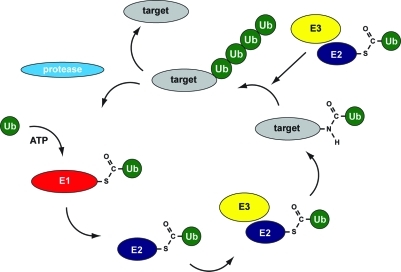
Ubiquitylation cascade. Ubiquitin precursors are processed by proteases to generate mature ubiquitin containing a C-terminal diglycine motif for conjugation to target proteins. Three different classes of enzymes are involved: E1, E2, and E3 enzymes. Ubiquitin is coupled to the active site cysteine of the E1 enzyme in an adenosine 5′-triphosphate-dependent manner to form a thioester. Subsequently, transfer to the active site cysteine in an E2 enzyme occurs, and a novel thioester is formed. With the help of E3 enzymes, ubiquitin is coupled to lysines in target proteins via isopeptide bonds, and in a subsequent optional step, ubiquitin chains can be formed. Specific proteases remove ubiquitin from target proteins, and free ubiquitin becomes available for novel rounds of conjugation.
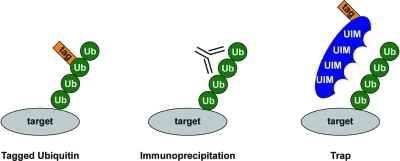
Ubiquitin purification strategies. Three strategies are available to purify ubiquitin. The first strategy employs epitope-tagged ubiquitin that is attached to target proteins. These tags are used to purify ubiquitin. The second strategy makes use of anti-ubiquitin antibodies to purify endogenous ubiquitin. The third strategy uses ubiquitin traps that consist of epitope-tagged proteins containing one or more ubiquitin interaction motifs (UIMs) to enrich for endogenous ubiquitin.
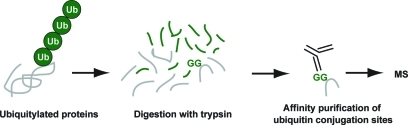
Site-specific ubiquitin purification methodology. A monoclonal antibody is available that recognizes diglycine motifs attached to lysines in target proteins. These diglycines become available upon digestion of ubiquitin conjugates with trypsin. After digestion, ubiquitylation sites are purified by immunoprecipitation and analyzed by MS. Note that the C-terminal tryptic peptides from ubiquitin and the UBLs Nedd8 and ISG15 are identical and that a pre-purification step is therefore required.
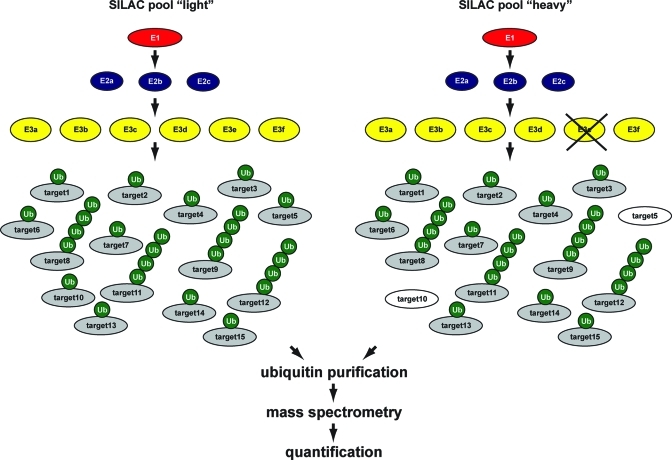
Quantitative proteomics strategy to identify target proteins for ubiquitin E3 enzymes. SILAC technology is used for differential labeling of cells to generate two distinct pools. Using RNAi or specific inhibitors, E3 enzymes can be inactivated. Target proteins that are regulated by the indicated E3 enzyme can be identified by purifying ubiquitin from both pools of cells and analyzing the purified samples by mass spectrometry. Decreases in SILAC ratios for specific proteins upon E3 inactivation will reveal potential target proteins for this E3 enzyme. Similar strategies can be used to identify target proteins for ubiquitin proteases or ubiquitin proteome dynamics in response to specific stimuli. A third SILAC pool can be added as a negative control. Alternative quantitative proteomics methodologies are also available.
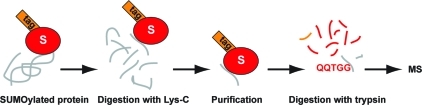
Site-specific SUMO purification methodology. An epitope-tagged lysine-deficient SUMO mutant can be used to purify and identify SUMOylation sites. Using this methodology, SUMO conjugates are digested with endopeptidase Lys-C, which cleaves C-terminal of lysine residues. After digestion, SUMO target proteins are cleaved, but the SUMO mutant stays intact and can be purified using the tag. The purified sample is subsequently digested with trypsin, which cleaves C-terminal of arginine residues in the SUMO mutant and in the target protein fragments. Introducing an arginine at the C-terminus of SUMO will provide convenient “QQTGG” SUMO tryptic fragments that can be identified by mass spectrometry. “QQ” ions are generated upon fragmentation of these purified SUMO sites and are useful as reporter ions. One example of these reporter ions is provided in Figure 6.

Example of a SUMOylation site identified by mass spectrometry. Lysine 497 of nucleolar protein NOP5/58 was identified as a SUMO-2 acceptor site using the SUMO-2 mutant Q87R. The identified tryptic fragment contained a phosphorylated serine on position 502. The doubly modified peptide was identified by high-resolution tandem MS using higher energy collisional dissociation. Inset: Magnification of the low mass region showing the “QQ” signature fragment ions. Reprinted with permission from ref (157). Copyright 2010 Elsevier.

SUMOylation consensus site. Graphical representation of the local target protein context of 103 SUMO-2-conjugated lysines that were identified using the method described in Figure 5. The SUMO-2 acceptor lysines were aligned using WebLogo. SUMOylation sites are frequently located in the consensus motif [VILMFPC]KxE. SUMOylation consensus motifs furthermore include the inverted SUMOylation consensus motif [ED]xK[VILFP] and HCSM. Reprinted with permission from ref (157). Copyright 2010 Elsevier.
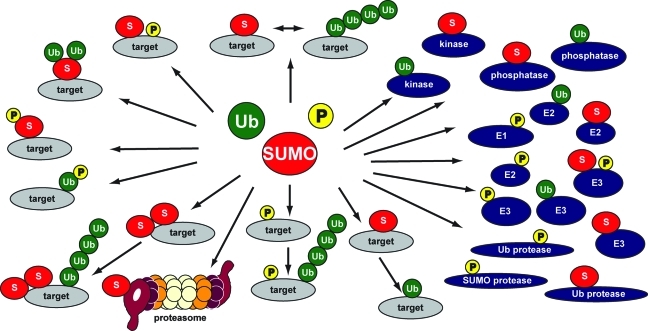
Crosstalk among ubiquitylation, SUMOylation, and phosphorylation. Extensive crosstalk among ubiquitylation, SUMOylation, and phosphorylation includes the modification of enzymatic components of the cascades. Furthermore, co-modification occurs along with competition for the same lysine residues in target proteins by SUMO and ubiquitin or consecutive modification by SUMO and ubiquitin of the same lysine residues. Moreover, noncovalent binding proteins add to the complexity but are not depicted here. Several examples are described in the main text.
References
-
- Garavelli J. S. Proteomics 2004, 4, 1527. - PubMed
-
- Aebersold R.; Goodlett D. R. Chem. Rev. 2001, 101, 269. - PubMed
-
- Choudhary C.; Mann M. Nat. Rev. Mol. Cell Biol. 2010, 11, 427. - PubMed
-
- Ding H.; Xu Y.; Chen Q.; Dai H.; Tang Y.; Wu J.; Shi Y. Biochemistry 2005, 44, 2790. - PubMed
-
- Bayer P.; Arndt A.; Metzger S.; Mahajan R.; Melchior F.; Jaenicke R.; Becker J. J. Mol. Biol. 1998, 280, 275. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
