

SREBP-dependent triazole susceptibility in Aspergillus fumigatus is mediated through direct transcriptional regulation of erg11A (cyp51A)
- PMID: 22006005
- PMCID: PMC3256050
- DOI: 10.1128/AAC.05027-11
SREBP-dependent triazole susceptibility in Aspergillus fumigatus is mediated through direct transcriptional regulation of erg11A (cyp51A)
Abstract
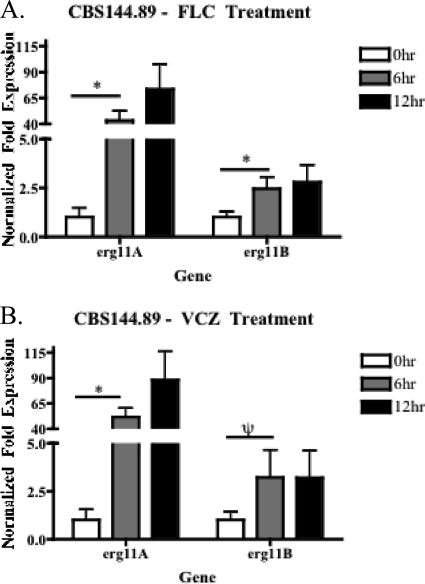
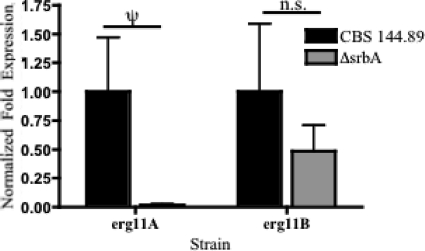
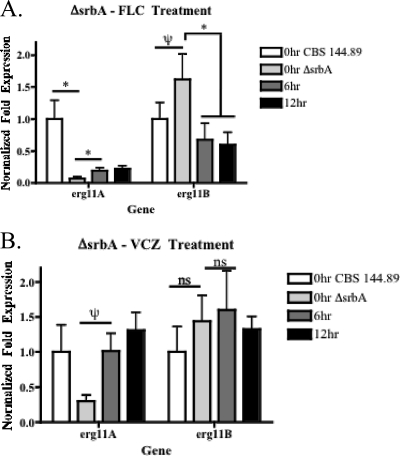
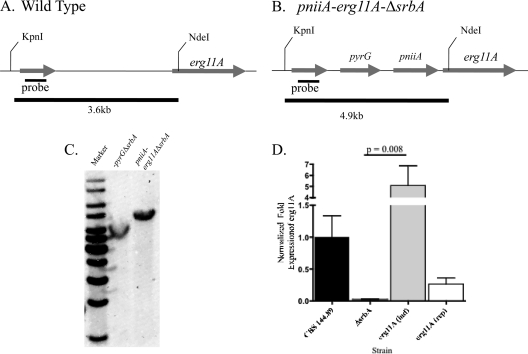
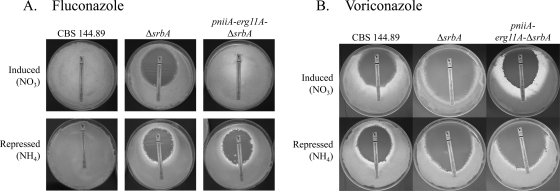
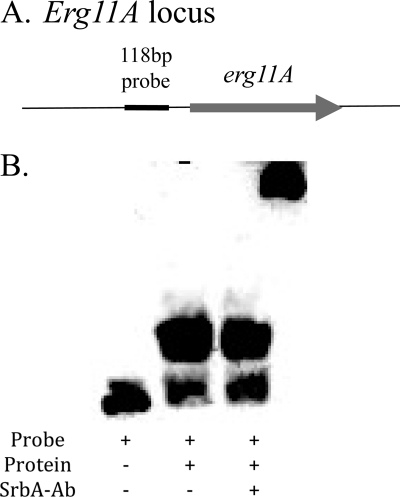
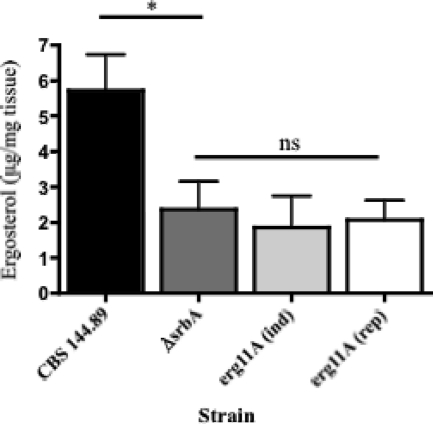
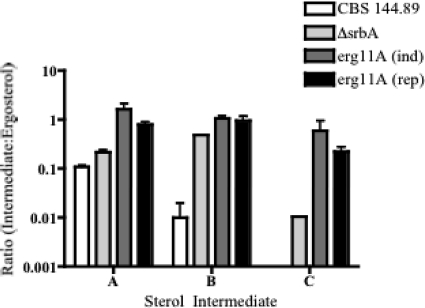
As triazole antifungal drug resistance during invasive Aspergillus fumigatus infection has become more prevalent, the need to understand mechanisms of resistance in A. fumigatus has increased. The presence of two erg11 (cyp51) genes in Aspergillus spp. is hypothesized to account for the inherent resistance of this mold to the triazole fluconazole (FLC). Recently, an A. fumigatus null mutant of a transcriptional regulator in the sterol regulatory element binding protein (SREBP) family, the ΔsrbA strain, was found to have increased susceptibility to FLC and voriconazole (VCZ). In this study, we examined the mechanism engendering the observed increase in A. fumigatus triazole susceptibility in the absence of SrbA. We observed a significant reduction in the erg11A transcript in the ΔsrbA strain in response to FLC and VCZ. Transcript levels of erg11B were also reduced but not to the extent of erg11A. Interestingly, erg11A transcript levels increased upon extended VCZ, but not FLC, exposure. Construction of an erg11A conditional expression strain in the ΔsrbA strain was able to restore erg11A transcript levels and, consequently, wild-type MICs to the triazole FLC. The VCZ MIC was also partially restored upon increased erg11A transcript levels; however, total ergosterol levels remained significantly reduced compared to those of the wild type. Induction of the erg11A conditional strain did not restore the hypoxia growth defect of the ΔsrbA strain. Taken together, our results demonstrate a critical role for SrbA-mediated regulation of ergosterol biosynthesis and triazole drug interactions in A. fumigatus that may have clinical importance.
Figures


References
-
- Albarrag AM, et al. 2011. Interrogation of related clinical pan-azole-resistant Aspergillus fumigatus strains: G138C, Y431C, and G434C single nucleotide polymorphisms in cyp51A, upregulation of cyp51A, and integration and activation of transposon Atf1 in the cyp51A promoter. Antimicrob. Agents Chemother. 55:5113–5121 - PMC - PubMed
-
- Alcazar-Fuoli L, Mellado E, Cuenca-Estrella M, Sanglard D. 2011. Probing the role of point mutations in the cyp51A gene from Aspergillus fumigatus in the model yeast Saccharomyces cerevisiae. Med. Mycol. 49:276–284 - PubMed
-
- Alcazar-Fuoli L, et al. 2008. Ergosterol biosynthesis pathway in Aspergillus fumigatus. Steroids 73:339–347 - PubMed
-
- Alexander BD, Perfect JR. 1997. Antifungal resistance trends towards the year 2000. Implications for therapy and new approaches. Drugs 54:657–678 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
