

Spatial constraints dictate glial territories at murine neuromuscular junctions
- PMID: 22006952
- PMCID: PMC3198169
- DOI: 10.1083/jcb.201108005
Spatial constraints dictate glial territories at murine neuromuscular junctions
Abstract
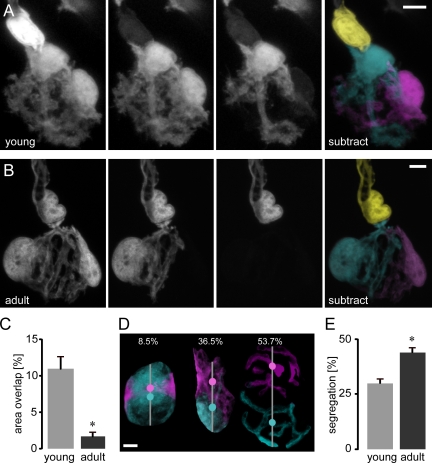
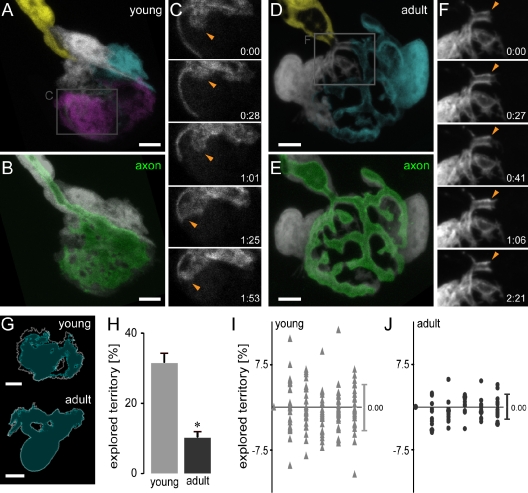
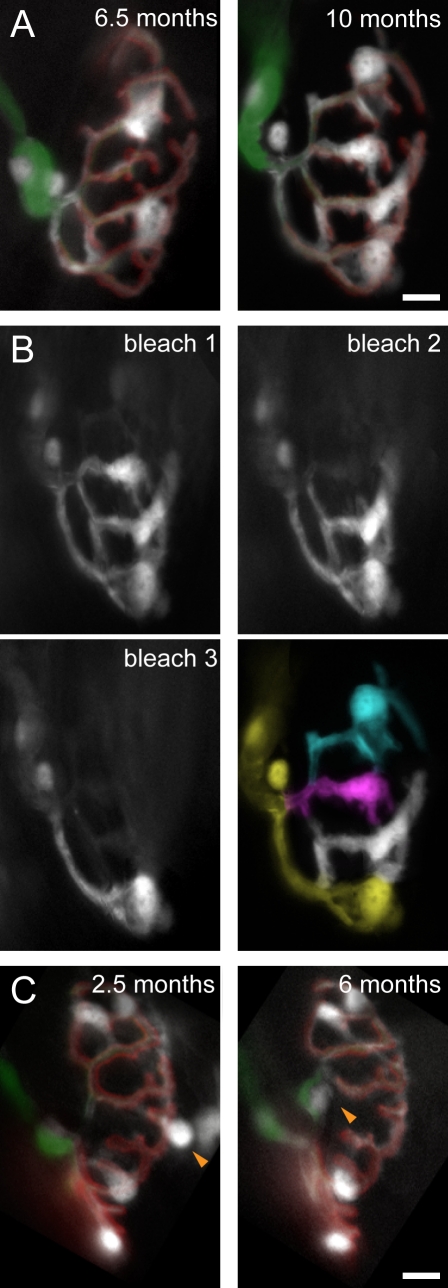
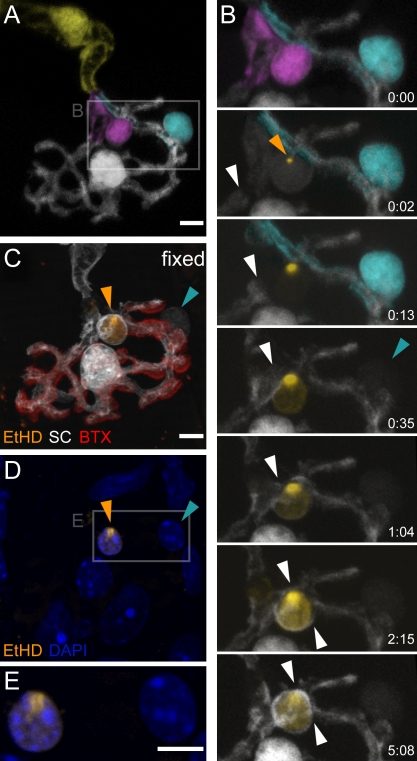
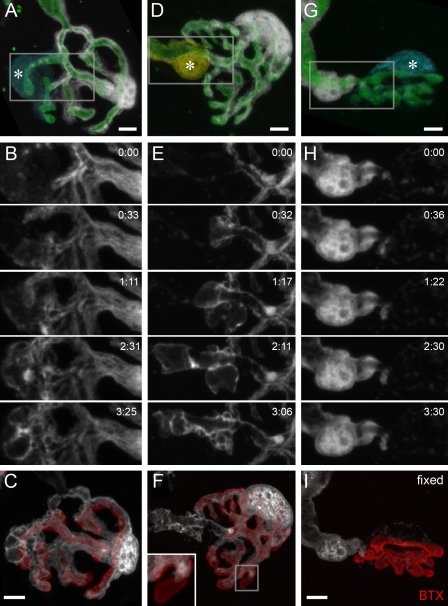
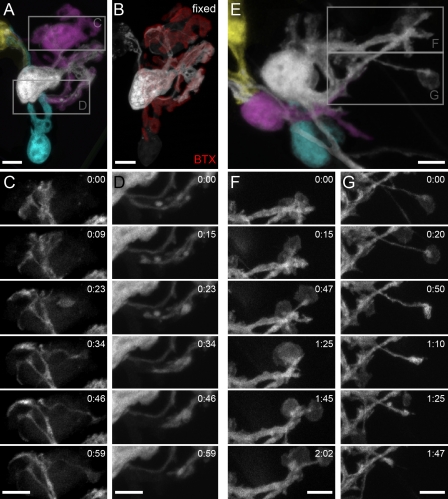
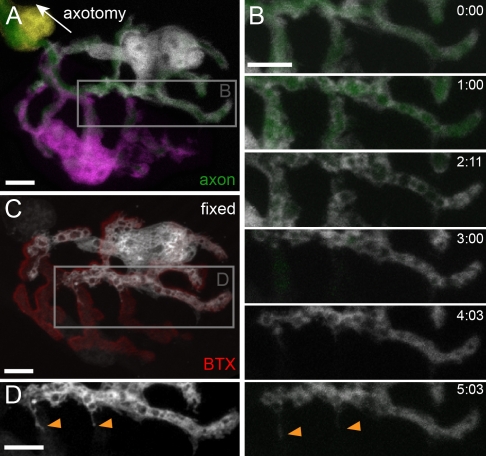
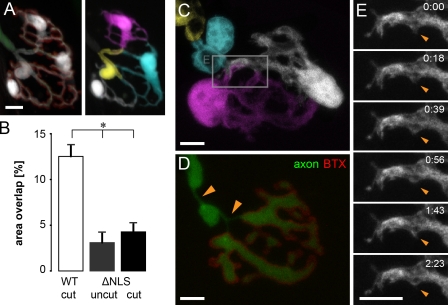
Schwann cells (SCs), the glial cells of the peripheral nervous system, cover synaptic terminals, allowing them to monitor and modulate neurotransmission. Disruption of glial coverage leads to axon degeneration and synapse loss. The cellular mechanisms that establish and maintain this coverage remain largely unknown. To address this, we labeled single SCs and performed time-lapse imaging experiments. Adult terminal SCs are arranged in static tile patterns, whereas young SCs dynamically intermingle. The mechanism of developmental glial segregation appears to be spatial competition, in which glial-glial and axonal-glial contacts constrain the territory of single SCs, as shown by four types of experiments: (1) laser ablation of single SCs, which led to immediate territory expansion of neighboring SCs; (2) axon removal by transection, resulting in adult SCs intermingling dynamically; (3) axotomy in mutant mice with blocked axon fragmentation in which intermingling was delayed; and (4) activity blockade, which had no immediate effects. In summary, we conclude that glial cells partition synapses by competing for perisynaptic space.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
