

Molecular dynamics simulations of the Cx26 hemichannel: evaluation of structural models with Brownian dynamics
- PMID: 22006989
- PMCID: PMC3206306
- DOI: 10.1085/jgp.201110679
Molecular dynamics simulations of the Cx26 hemichannel: evaluation of structural models with Brownian dynamics
Abstract
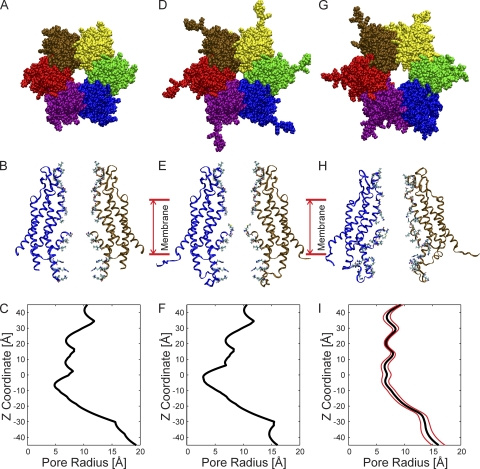
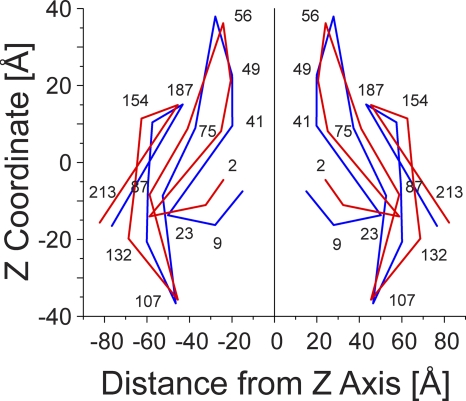
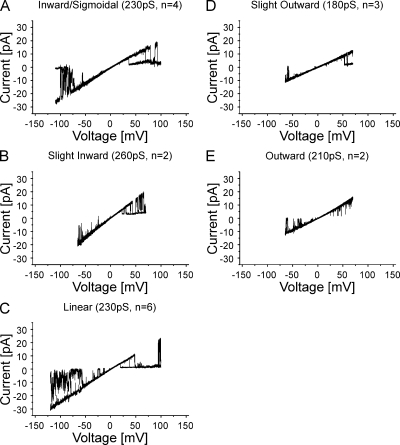
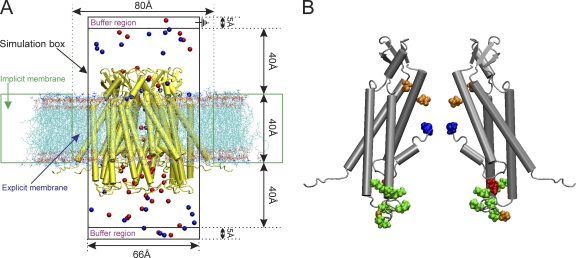
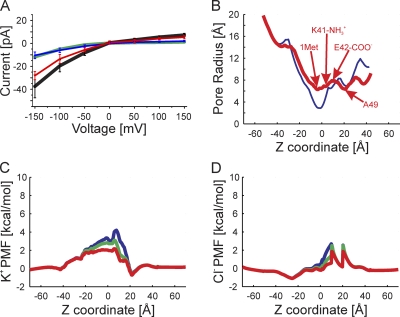
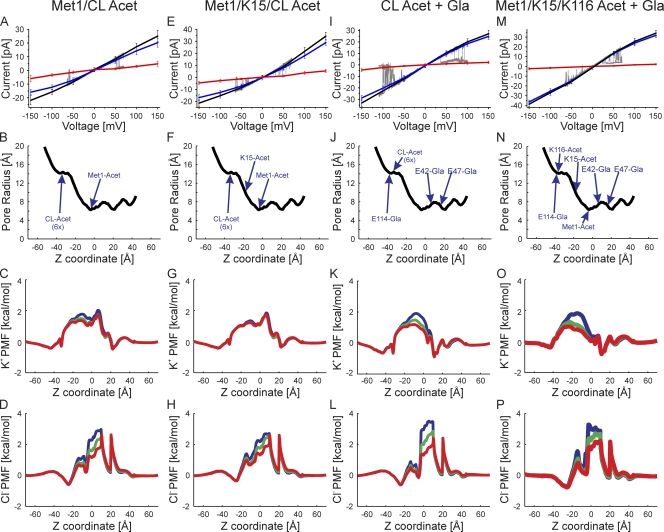
The recently published crystal structure of the Cx26 gap junction channel provides a unique opportunity for elucidation of the structure of the conductive connexin pore and the molecular determinants of its ion permeation properties (conductance, current-voltage [I-V] relations, and charge selectivity). However, the crystal structure was incomplete, most notably lacking the coordinates of the N-terminal methionine residue, which resides within the pore, and also lacking two cytosolic domains. To allow computational studies for comparison with the known channel properties, we completed the structure. Grand canonical Monte Carlo Brownian dynamics (GCMC/BD) simulations of the completed and the published Cx26 hemichannel crystal structure indicate that the pore is too narrow to permit significant ion flux. The GCMC/BD simulations predict marked inward current rectification and almost perfect anion selectivity, both inconsistent with known channel properties. The completed structure was refined by all-atom molecular dynamics (MD) simulations (220 ns total) in an explicit solvent and POPC membrane system. These MD simulations produced an equilibrated structure with a larger minimal pore diameter, which decreased the height of the permeation barrier formed by the N terminus. GCMC/BD simulations of the MD-equilibrated structure yielded more appropriate single-channel conductance and less anion/cation selectivity. However, the simulations much more closely matched experimentally determined I-V relations when the charge effects of specific co- and posttranslational modifications of Cx26 previously identified by mass spectrometry were incorporated. We conclude that the average equilibrated structure obtained after MD simulations more closely represents the open Cx26 hemichannel structure than does the crystal structure, and that co- and posttranslational modifications of Cx26 hemichannels are likely to play an important physiological role by defining the conductance and ion selectivity of Cx26 channels. Furthermore, the simulations and data suggest that experimentally observed heterogeneity in Cx26 I-V relations can be accounted for by variation in co- and posttranslational modifications.
Figures
Similar articles
-
Voltage-dependent conformational changes in connexin channels.Biochim Biophys Acta. 2012 Aug;1818(8):1807-22. doi: 10.1016/j.bbamem.2011.09.019. Epub 2011 Sep 24. Biochim Biophys Acta. 2012. PMID: 21978595 Free PMC article. Review.
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Role of gamma carboxylated Glu47 in connexin 26 hemichannel regulation by extracellular Ca²⁺: insight from a local quantum chemistry study.Biochem Biophys Res Commun. 2014 Feb 28;445(1):10-5. doi: 10.1016/j.bbrc.2014.01.063. Epub 2014 Jan 24. Biochem Biophys Res Commun. 2014. PMID: 24468086 Free PMC article.
-
The D50N mutation and syndromic deafness: altered Cx26 hemichannel properties caused by effects on the pore and intersubunit interactions.J Gen Physiol. 2013 Jul;142(1):3-22. doi: 10.1085/jgp.201310962. J Gen Physiol. 2013. PMID: 23797419 Free PMC article.
-
Different ionic selectivities for connexins 26 and 32 produce rectifying gap junction channels.Biophys J. 1999 Dec;77(6):2968-87. doi: 10.1016/S0006-3495(99)77129-8. Biophys J. 1999. PMID: 10585920 Free PMC article.
Cited by
-
Specificity of the connexin W3/4 locus for functional gap junction formation.Channels (Austin). 2016 Nov;10(6):453-65. doi: 10.1080/19336950.2016.1200775. Epub 2016 Jun 15. Channels (Austin). 2016. PMID: 27304225 Free PMC article.
-
The N-terminal half of the connexin protein contains the core elements of the pore and voltage gates.J Membr Biol. 2012 Aug;245(8):453-63. doi: 10.1007/s00232-012-9457-z. Epub 2012 Jul 24. J Membr Biol. 2012. PMID: 22825713 Free PMC article.
-
Gap-junctional channel and hemichannel activity of two recently identified connexin 26 mutants associated with deafness.Pflugers Arch. 2016 May;468(5):909-18. doi: 10.1007/s00424-016-1788-7. Epub 2016 Jan 14. Pflugers Arch. 2016. PMID: 26769242
-
Gating of Connexin Channels by transjunctional-voltage: Conformations and models of open and closed states.Biochim Biophys Acta Biomembr. 2018 Jan;1860(1):22-39. doi: 10.1016/j.bbamem.2017.04.028. Epub 2017 May 2. Biochim Biophys Acta Biomembr. 2018. PMID: 28476631 Free PMC article. Review.
-
Keratitis-ichthyosis-deafness syndrome-associated Cx26 mutants produce nonfunctional gap junctions but hyperactive hemichannels when co-expressed with wild type Cx43.J Invest Dermatol. 2015 May;135(5):1338-1347. doi: 10.1038/jid.2015.20. Epub 2015 Jan 27. J Invest Dermatol. 2015. PMID: 25625422 Free PMC article.
References
-
- Allen M.J., Gemel J., Beyer E.C., Lal R. 2011. Atomic force microscopy of Connexin40 gap junction hemichannels reveals calcium-dependent three-dimensional molecular topography and open-closed conformations of both the extracellular and cytoplasmic faces. J. Biol. Chem. 286:22139–22146 10.1074/jbc.M111.240002 - DOI - PMC - PubMed
-
- Arnesen T., Van Damme P., Polevoda B., Helsens K., Evjenth R., Colaert N., Varhaug J.E., Vandekerckhove J., Lillehaug J.R., Sherman F., Gevaert K. 2009. Proteomics analyses reveal the evolutionary conservation and divergence of N-terminal acetyltransferases from yeast and humans. Proc. Natl. Acad. Sci. USA. 106:8157–8162 10.1073/pnas.0901931106 - DOI - PMC - PubMed
-
- Bargiello T., Brink P. 2009. Voltage-gating mechanisms of connexin channels. Connexins. Harris A.L., Locke D., Humana Press, New York: 103–128
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
