

HESX1- and TCF3-mediated repression of Wnt/β-catenin targets is required for normal development of the anterior forebrain
- PMID: 22007134
- PMCID: PMC3201662
- DOI: 10.1242/dev.066597
HESX1- and TCF3-mediated repression of Wnt/β-catenin targets is required for normal development of the anterior forebrain
Abstract
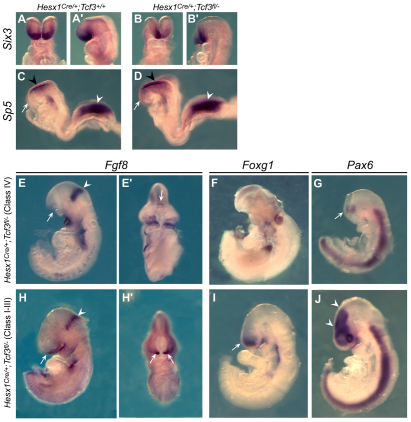
The Wnt/β-catenin pathway plays an essential role during regionalisation of the vertebrate neural plate and its inhibition in the most anterior neural ectoderm is required for normal forebrain development. Hesx1 is a conserved vertebrate-specific transcription factor that is required for forebrain development in Xenopus, mice and humans. Mouse embryos deficient for Hesx1 exhibit a variable degree of forebrain defects, but the molecular mechanisms underlying these defects are not fully understood. Here, we show that injection of a hesx1 morpholino into a 'sensitised' zygotic headless (tcf3) mutant background leads to severe forebrain and eye defects, suggesting an interaction between Hesx1 and the Wnt pathway during zebrafish forebrain development. Consistent with a requirement for Wnt signalling repression, we highlight a synergistic gene dosage-dependent interaction between Hesx1 and Tcf3, a transcriptional repressor of Wnt target genes, to maintain anterior forebrain identity during mouse embryogenesis. In addition, we reveal that Tcf3 is essential within the neural ectoderm to maintain anterior character and that its interaction with Hesx1 ensures the repression of Wnt targets in the developing forebrain. By employing a conditional loss-of-function approach in mouse, we demonstrate that deletion of β-catenin, and concomitant reduction of Wnt signalling in the developing anterior forebrain of Hesx1-deficient embryos, leads to a significant rescue of the forebrain defects. Finally, transcriptional profiling of anterior forebrain precursors from mouse embryos expressing eGFP from the Hesx1 locus provides molecular evidence supporting a novel function of Hesx1 in mediating repression of Wnt/β-catenin target activation in the developing forebrain.
Figures




References
-
- Brannon M., Brown J. D., Bates R., Kimelman D., Moon R. T. (1999). XCtBP is a XTcf-3 co-repressor with roles throughout Xenopus development. Development 126, 3159–3170 - PubMed
-
- Brault V., Moore R., Kutsch S., Ishibashi M., Rowitch D. H., McMahon A. P., Sommer L., Boussadia O., Kemler R. (2001). Inactivation of the beta-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development 128, 1253–1264 - PubMed
-
- Braun M. M., Etheridge A., Bernard A., Robertson C. P., Roelink H. (2003). Wnt signaling is required at distinct stages of development for the induction of the posterior forebrain. Development 130, 5579–5587 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
