

Proteomic divergence in Arabidopsis autopolyploids and allopolyploids and their progenitors
- PMID: 22009271
- PMCID: PMC3313054
- DOI: 10.1038/hdy.2011.92
Proteomic divergence in Arabidopsis autopolyploids and allopolyploids and their progenitors
Abstract
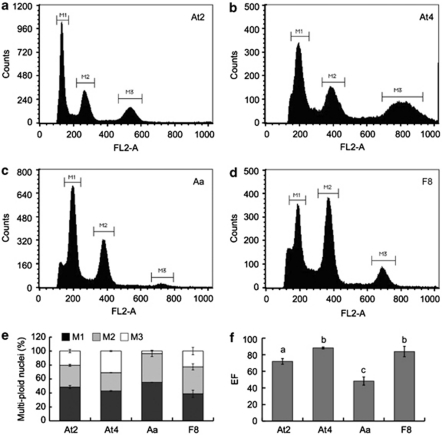
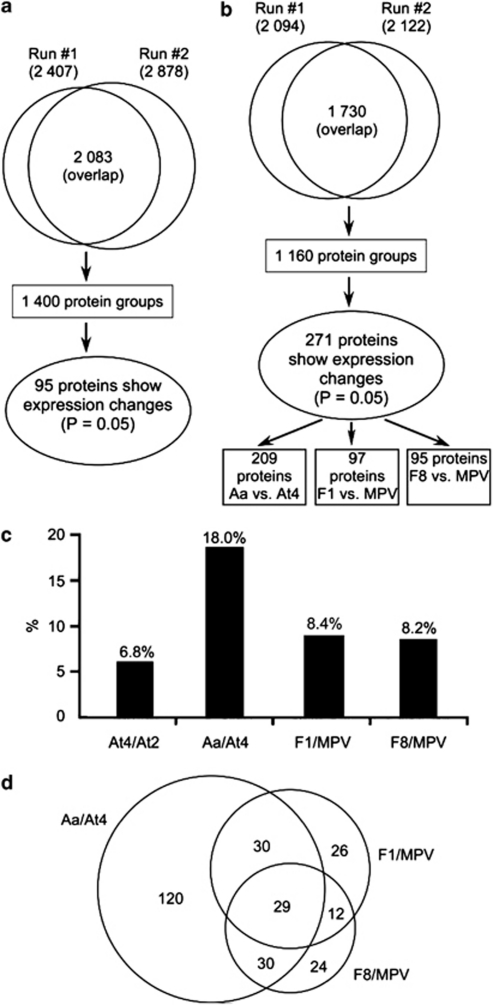
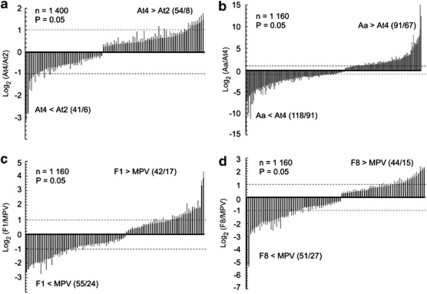
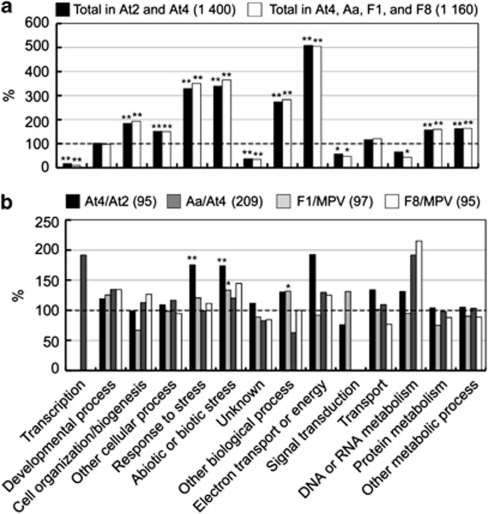
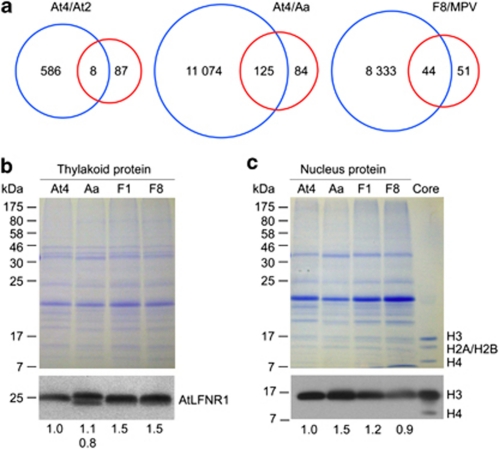
Autopolyploidy and allopolyploidy are common in many plants and some animals. Rapid changes in genomic composition and gene expression have been observed in both autopolyploids and allopolyploids, but the effects of polyploidy on proteomic divergence are poorly understood. Here, we report quantitative analysis of protein changes in leaves of Arabidopsis autopolyploids and allotetraploids and their progenitors using isobaric tags for relative and absolute quantitation (iTRAQ) coupled with mass spectrometry. In more than 1000 proteins analyzed, the levels of protein divergence were relatively high (~18%) between Arabidopsis thaliana and Arabidopsis arenosa, relatively low (~6.8%) between an A. thaliana diploid and autotetraploid and intermediate (~8.3 and 8.2%) in F(1)- and F(8)-resynthesized allotetraploids relative to mid-parent values, respectively. This pattern of proteomic divergence was consistent with the previously reported gene expression data. In particular, many non-additively accumulated proteins (61-62%) in the F(1) and F(8) allotetraploids were also differentially expressed between the parents. The differentially accumulated proteins in functional categories of abiotic and biotic stresses were overrepresented between an A. thaliana autotetraploid and diploid and between two Arabidopsis species, but not significantly different between allotetraploids and their progenitors. Although the trend of changes is similar, the percentage of differentially accumulated proteins that matched previously reported differentially expressed genes was relatively low. Western blot analysis confirmed several selected proteins with isoforms the cumulative levels of which were differentially expressed. These data suggest high protein divergence between species and rapid changes in post-transcriptional regulation and translational modifications of proteins during polyploidization.
Figures

References
-
- Albertin W, Brabant P, Catrice O, Eber F, Jenczewski E, Chevre AM, et al. Autopolyploidy in cabbage (Brassica oleracea L.) does not alter significantly the proteomes of green tissues. Proteomics. 2005;5:2131–2139. - PubMed
-
- Albertin W, Langella O, Joets J, Negroni L, Zivy M, Damerval C, et al. Comparative proteomics of leaf, stem, and root tissues of synthetic Brassica napus. Proteomics. 2009;9:793–799. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
