

The composition of plant mitochondrial supercomplexes changes with oxygen availability
- PMID: 22009743
- PMCID: PMC3234806
- DOI: 10.1074/jbc.M111.252544
The composition of plant mitochondrial supercomplexes changes with oxygen availability
Abstract
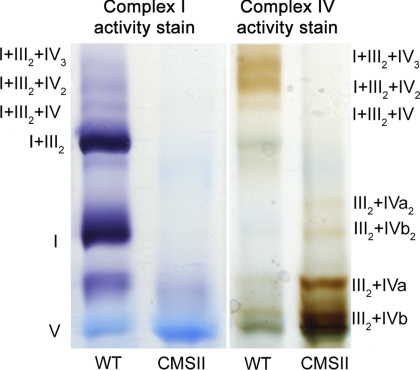
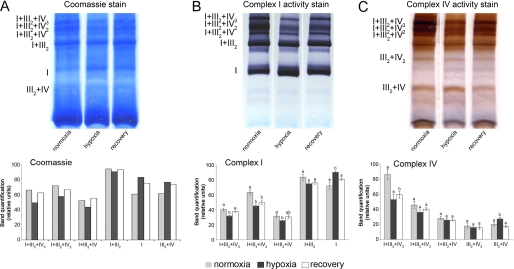
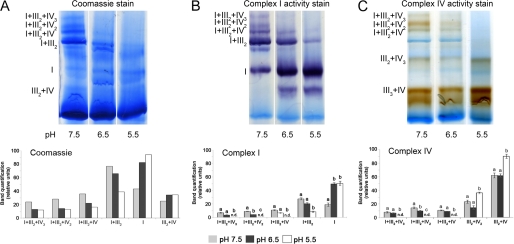
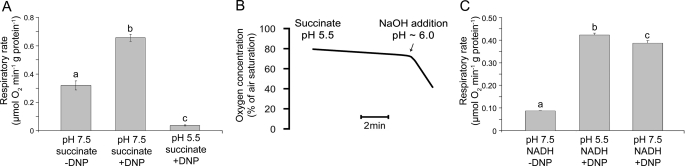
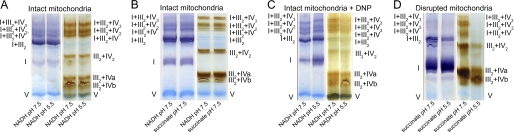
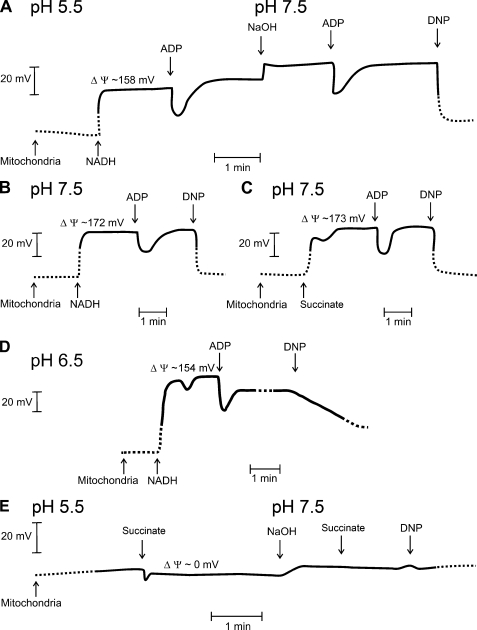
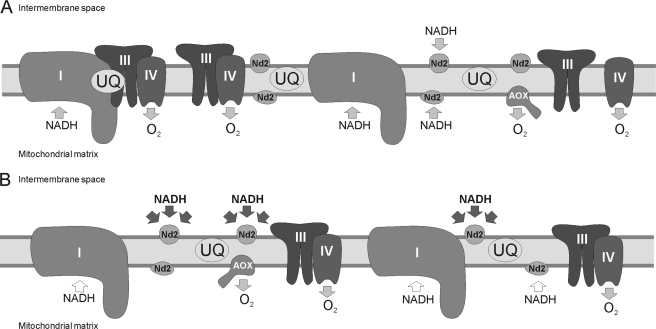
Respiratory supercomplexes are large protein structures formed by various enzyme complexes of the mitochondrial electron transport chain. Using native gel electrophoresis and activity staining, differential regulation of complex activity within the supercomplexes was investigated. During prolonged hypoxia, complex I activity within supercomplexes diminished, whereas the activity of the individual complex I-monomer increased. Concomitantly, an increased activity was observed during hypoxia for complex IV in the smaller supercomplexes that do not contain complex I. These changes in complex activity within supercomplexes reverted again during recovery from the hypoxic treatment. Acidification of the mitochondrial matrix induced similar changes in complex activity within the supercomplexes. It is suggested that the increased activity of the small supercomplex III(2)+IV can be explained by the dissociation of complex I from the large supercomplexes. This is discussed to be part of a mechanism regulating the involvement of the alternative NADH dehydrogenases, known to be activated by low pH, and complex I, which is inhibited by low pH. It is concluded that the activity of complexes within supercomplexes can be regulated depending on the oxygen status and the pH of the mitochondrial matrix.
Figures
References
-
- Schäfer E., Dencher N. A., Vonck J., Parcej D. N. (2007) Biochemistry 46, 12579–12585 - PubMed
-
- Eubel H., Heinemeyer J., Sunderhaus S., Braun H. P. (2004) Plant Physiol. Biochem. 42, 937–942 - PubMed
-
- Bultema J. B., Braun H. P., Boekema E. J., Kouril R. (2009) Biochim. Biophys. Acta 1787, 60–67 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
