

Deletion of the de novo DNA methyltransferase Dnmt3a promotes lung tumor progression
- PMID: 22011581
- PMCID: PMC3207684
- DOI: 10.1073/pnas.1114946108
Deletion of the de novo DNA methyltransferase Dnmt3a promotes lung tumor progression
Abstract
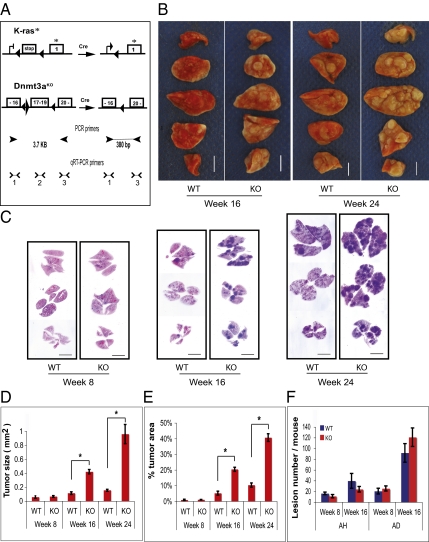
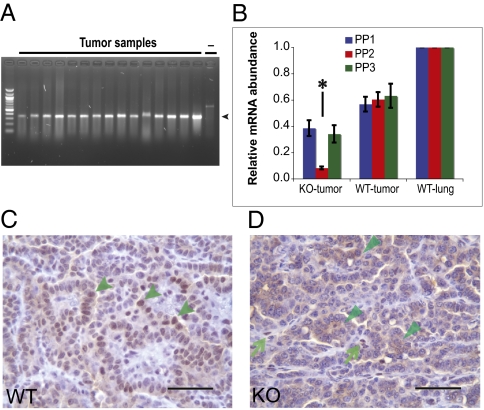
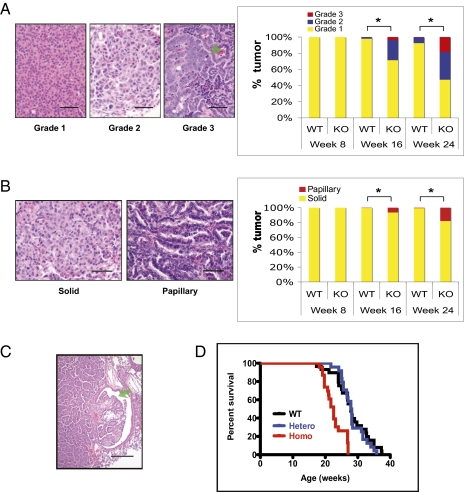
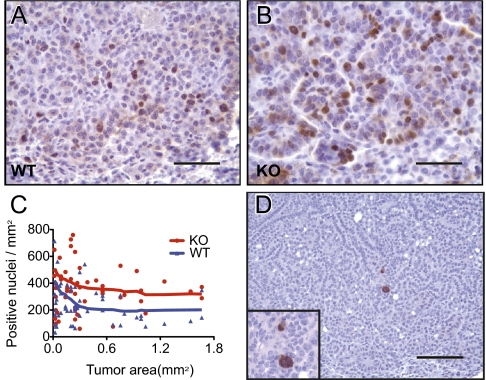
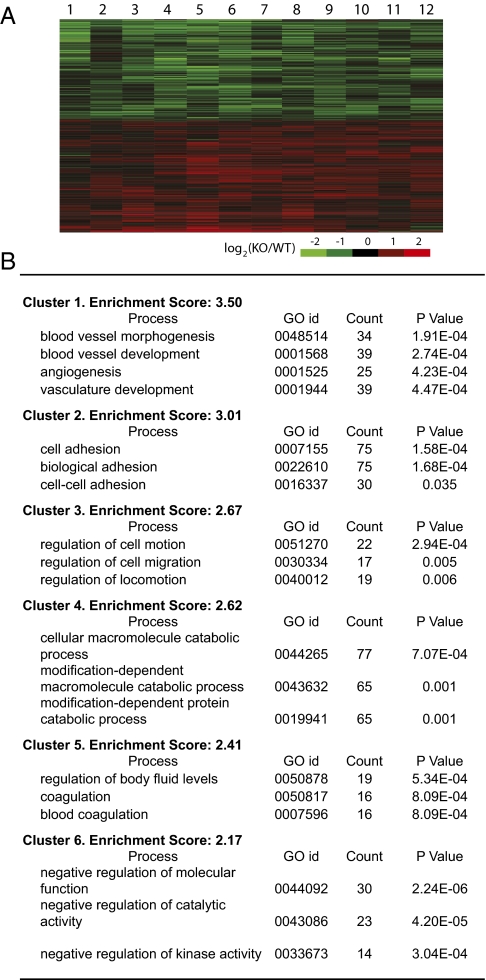
Alterations in DNA methylation have been associated with genome-wide hypomethylation and regional de novo methylation in numerous cancers. De novo methylation is mediated by the de novo methyltransferases Dnmt3a and 3b, but only Dnmt3b has been implicated in promoting cancer by silencing of tumor-suppressor genes. In this study, we have analyzed the role of Dnmt3a in lung cancer by using a conditional mouse tumor model. We show that Dnmt3a deficiency significantly promotes tumor growth and progression but not initiation. Changes in gene expression show that Dnmt3a deficiency affects key steps in cancer progression, such as angiogenesis, cell adhesion, and cell motion, consistent with accelerated and more malignant growth. Our results suggest that Dnmt3a may act like a tumor-suppressor gene in lung tumor progression and may be a critical determinant of lung cancer malignancy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Feinberg AP. Phenotypic plasticity and the epigenetics of human disease. Nature. 2007;447:433–440. - PubMed
-
- Gaudet F, et al. Induction of tumors in mice by genomic hypomethylation. Science. 2003;300:489–492. - PubMed
-
- Chen RZ, Pettersson U, Beard C, Jackson-Grusby L, Jaenisch R. DNA hypomethylation leads to elevated mutation rates. Nature. 1998;395:89–93. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
