

Axonemal positioning and orientation in three-dimensional space for primary cilia: what is known, what is assumed, and what needs clarification
- PMID: 22012592
- PMCID: PMC3278774
- DOI: 10.1002/dvdy.22756
Axonemal positioning and orientation in three-dimensional space for primary cilia: what is known, what is assumed, and what needs clarification
Abstract
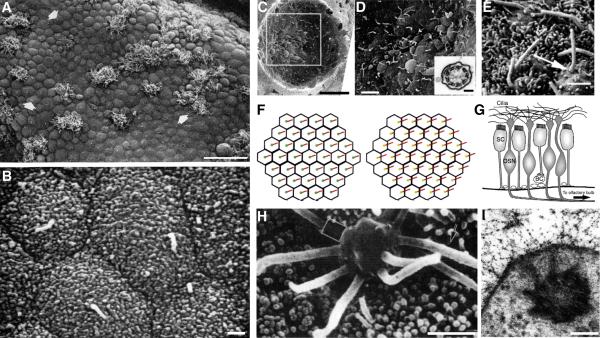
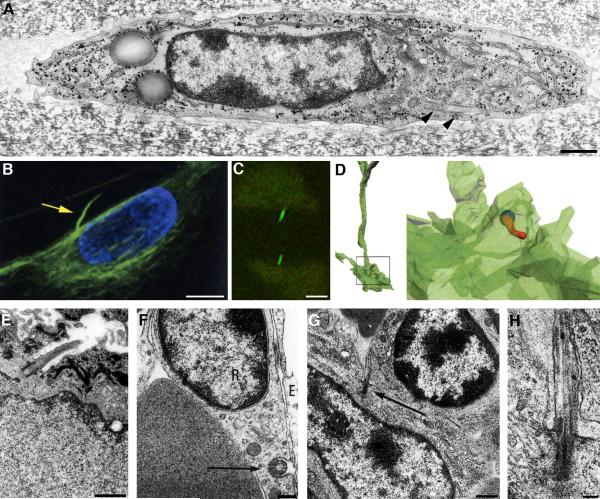
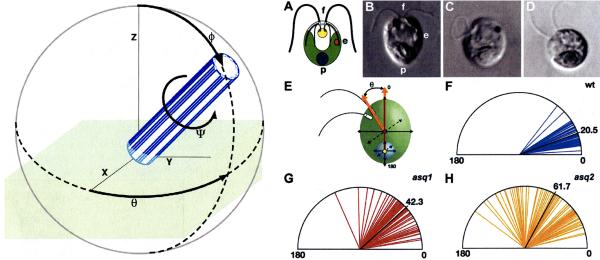
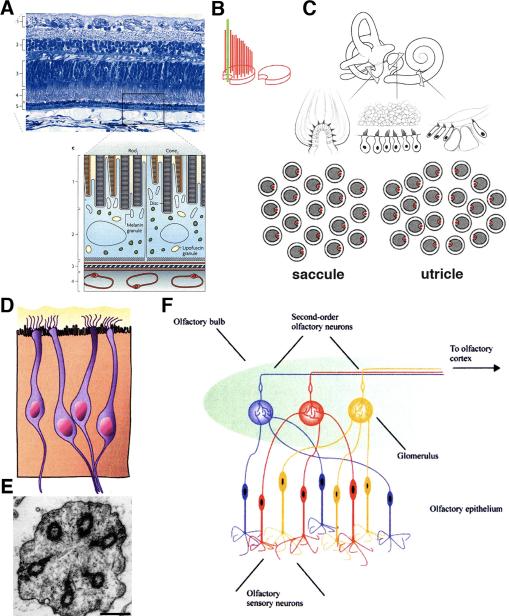
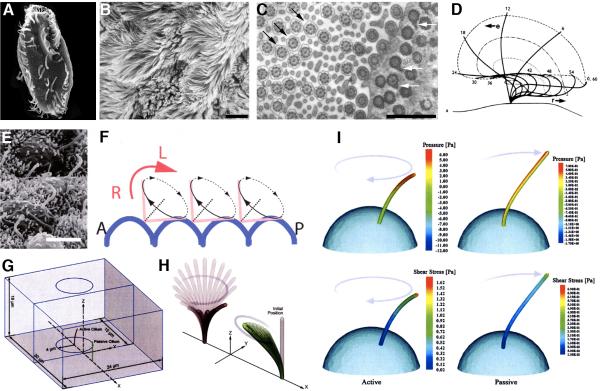
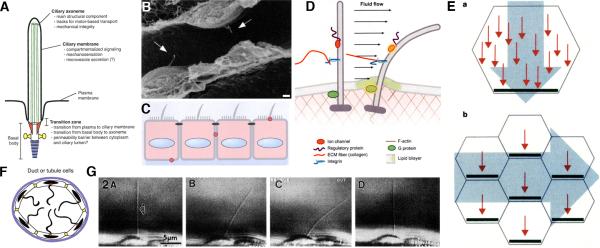
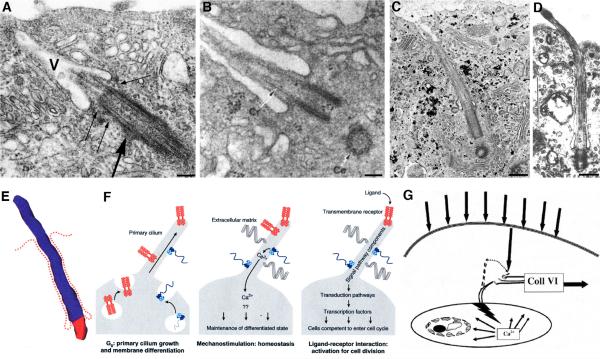
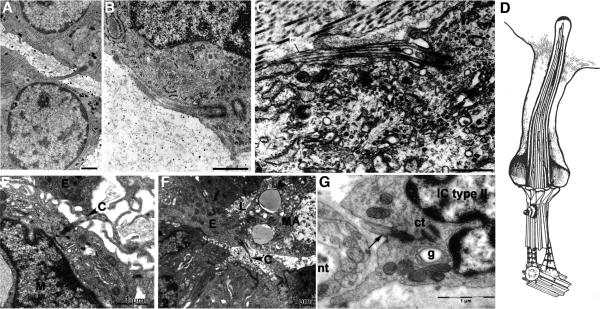
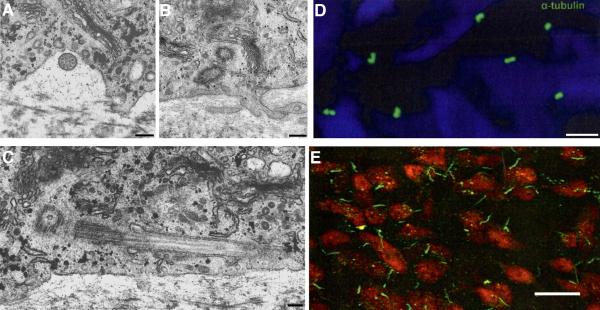
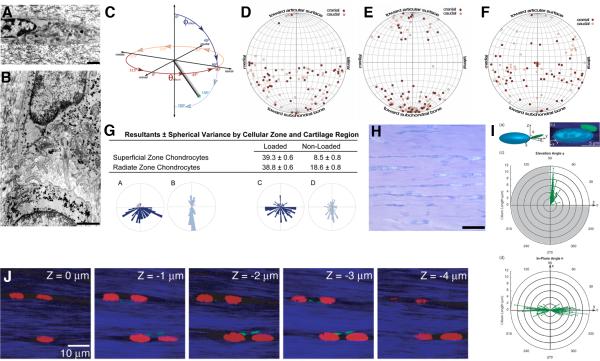
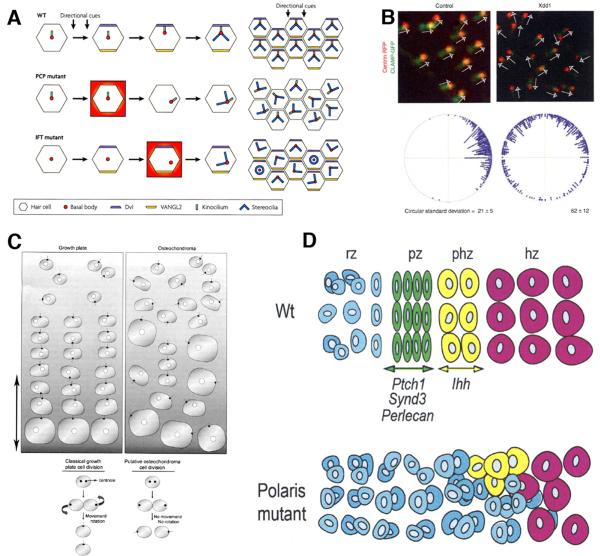
Two positional characteristics of the ciliary axoneme--its location on the plasma membrane as it emerges from the cell, and its orientation in three-dimensional (3D) space--are known to be critical for optimal function of actively motile cilia (including nodal cilia), as well as for modified cilia associated with special senses. However, these positional characteristics have not been analyzed to any significant extent for primary cilia. This review briefly summarizes the history of knowledge of these two positional characteristics across a wide spectrum of cilia, emphasizing their importance for proper function. Then the review focuses what is known about these same positional characteristics for primary cilia in all major tissue types where they have been reported. The review emphasizes major areas that would be productive for future research for understanding how positioning and 3D orientation of primary cilia may be related to their hypothesized signaling roles within different cellular populations.
Copyright © 2011 Wiley Periodicals, Inc.
Figures
References
-
- Afzelius BA. The immotile-cilia syndrome: a microtubule-associated defect. CRC Crit Rev Biochem. 1985;19:63–87. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
