

Control of blood glucose in the absence of hepatic glucose production during prolonged fasting in mice: induction of renal and intestinal gluconeogenesis by glucagon
- PMID: 22013018
- PMCID: PMC3219939
- DOI: 10.2337/db11-0571
Control of blood glucose in the absence of hepatic glucose production during prolonged fasting in mice: induction of renal and intestinal gluconeogenesis by glucagon
Abstract
Objective: Since the pioneering work of Claude Bernard, the scientific community has considered the liver to be the major source of endogenous glucose production in all postabsorptive situations. Nevertheless, the kidneys and intestine can also produce glucose in blood, particularly during fasting and under protein feeding. The aim of this study was to better define the importance of the three gluconeogenic organs in glucose homeostasis.
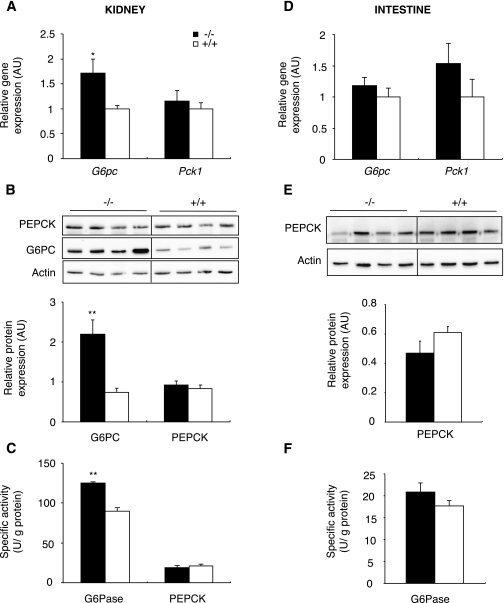
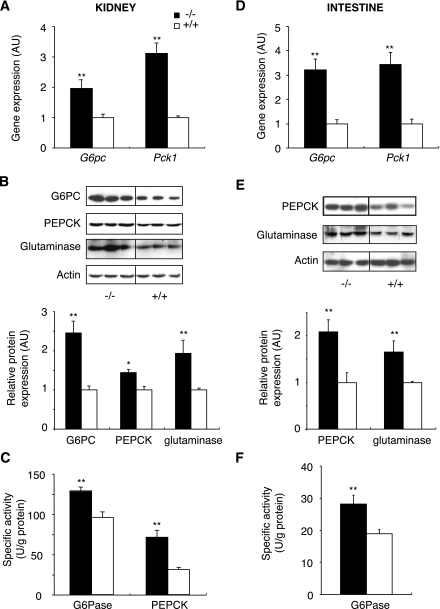
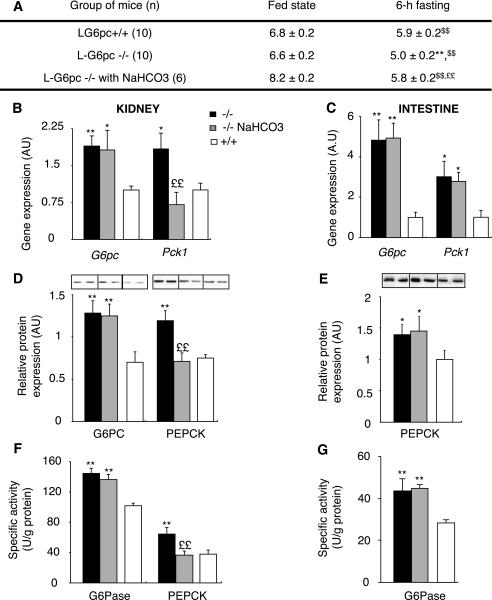
Research design and methods: We investigated blood glucose regulation during fasting in a mouse model of inducible liver-specific deletion of the glucose-6-phosphatase gene (L-G6pc(-/-) mice), encoding a mandatory enzyme for glucose production. Furthermore, we characterized molecular mechanisms underlying expression changes of gluconeogenic genes (G6pc, Pck1, and glutaminase) in both the kidneys and intestine.
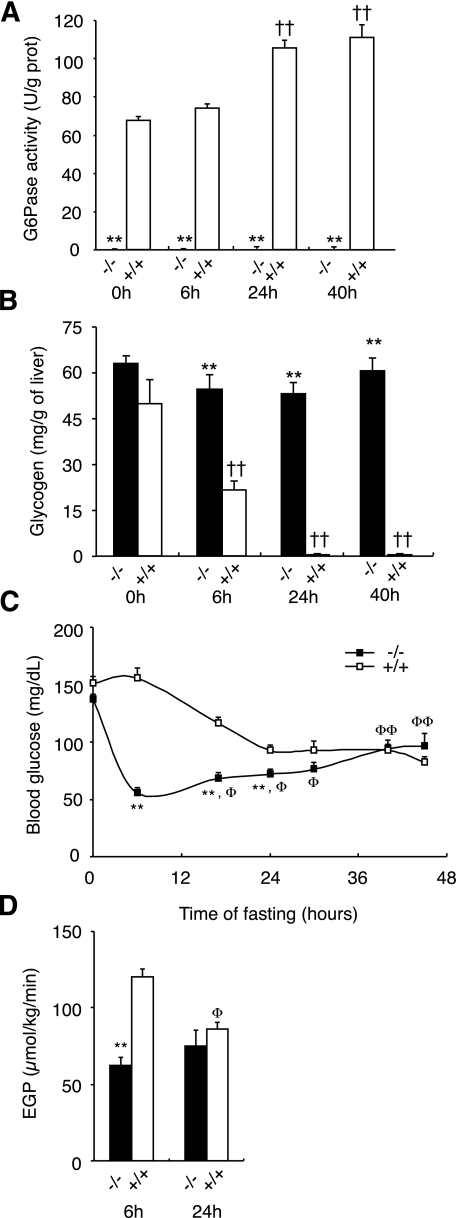
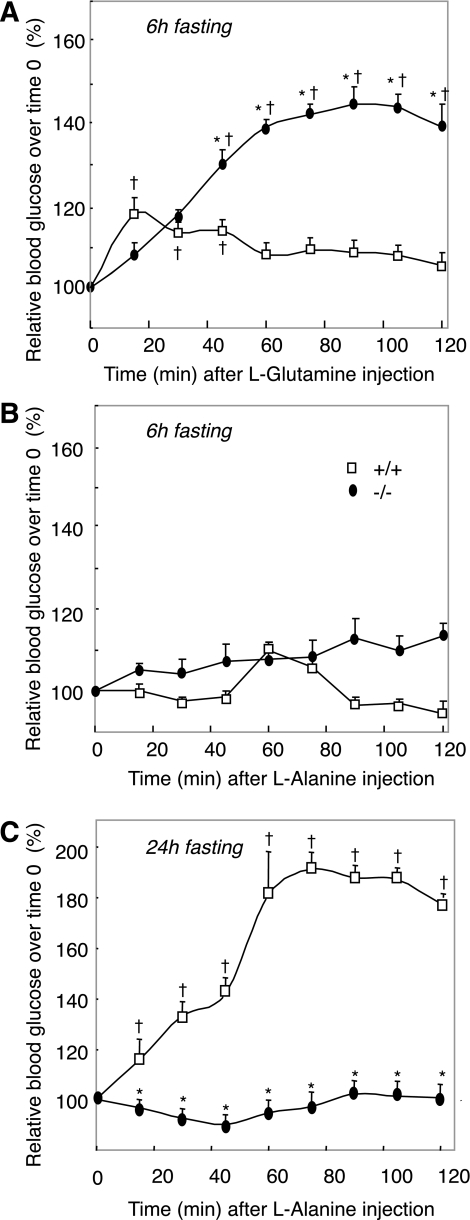
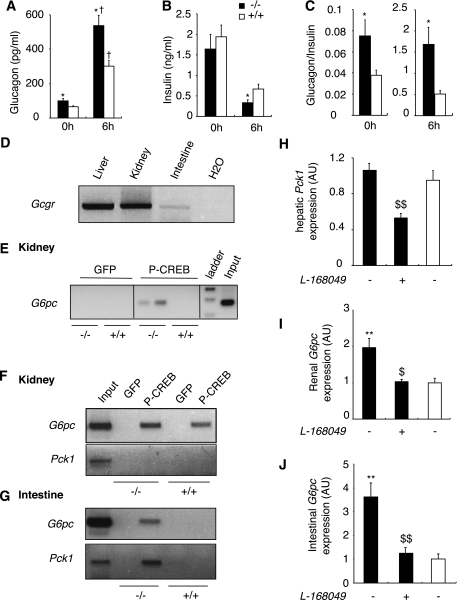
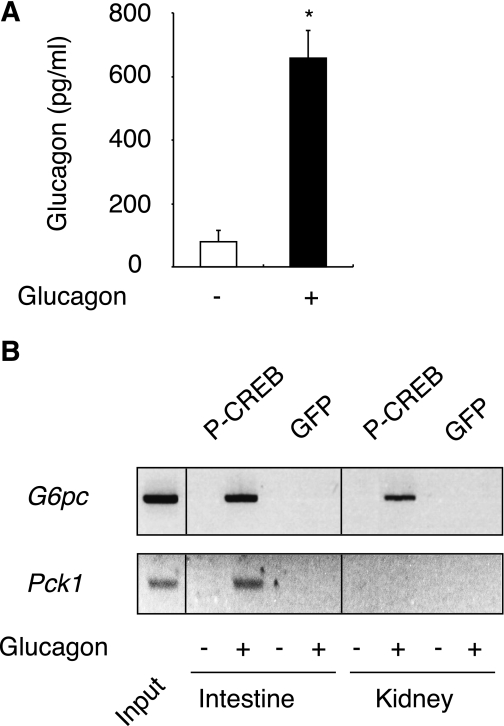
Results: We show that the absence of hepatic glucose release had no major effect on the control of fasting plasma glucose concentration. Instead, compensatory induction of gluconeogenesis occurred in the kidneys and intestine, driven by glucagon, glucocorticoids, and acidosis. Moreover, the extrahepatic action of glucagon took place in wild-type mice.
Conclusions: Our study provides a definitive quantitative estimate of the capacity of extrahepatic gluconeogenesis to sustain fasting endogenous glucose production under the control of glucagon, regardless of the contribution of the liver. Thus, the current dogma relating to the respective role of the liver and of extrahepatic gluconeogenic organs in glucose homeostasis requires re-examination.
Figures
References
-
- Wahren J, Ekberg K. Splanchnic regulation of glucose production. Annu Rev Nutr 2007;27:329–345 - PubMed
-
- Croset M, Rajas F, Zitoun C, Hurot JM, Montano S, Mithieux G. Rat small intestine is an insulin-sensitive gluconeogenic organ. Diabetes 2001;50:740–746 - PubMed
-
- Ekberg K, Landau BR, Wajngot A, et al. Contributions by kidney and liver to glucose production in the postabsorptive state and after 60 h of fasting. Diabetes 1999;48:292–298 - PubMed
-
- Gerich JE, Meyer C, Woerle HJ, Stumvoll M. Renal gluconeogenesis: its importance in human glucose homeostasis. Diabetes Care 2001;24:382–391 - PubMed
-
- Mithieux G, Bady I, Gautier A, Croset M, Rajas F, Zitoun C. Induction of control genes in intestinal gluconeogenesis is sequential during fasting and maximal in diabetes. Am J Physiol Endocrinol Metab 2004;286:E370–E375 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
