

The DNA damage response induces IFN
- PMID: 22013119
- PMCID: PMC3246365
- DOI: 10.4049/jimmunol.1100040
The DNA damage response induces IFN
Abstract
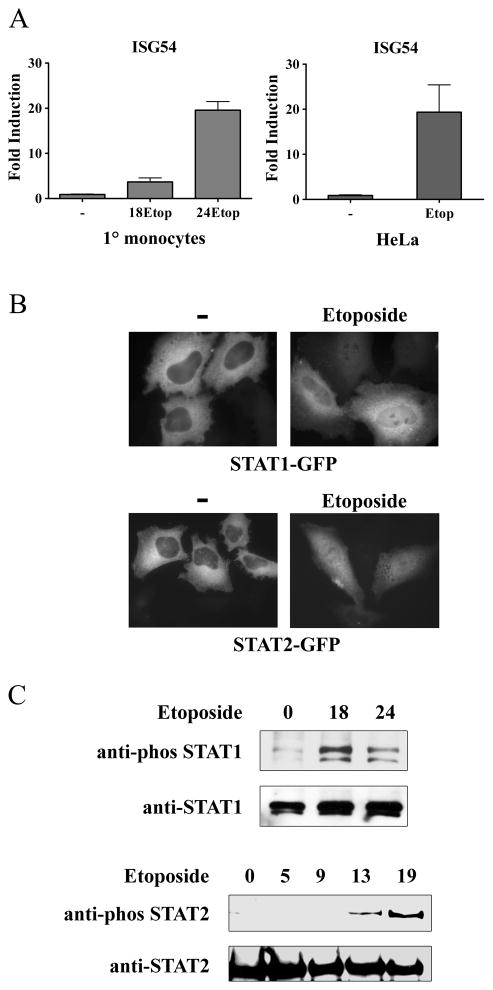
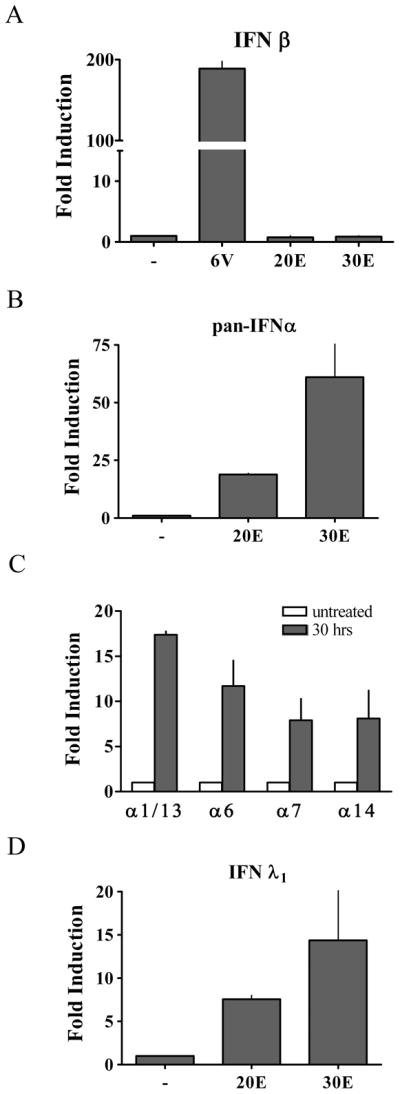
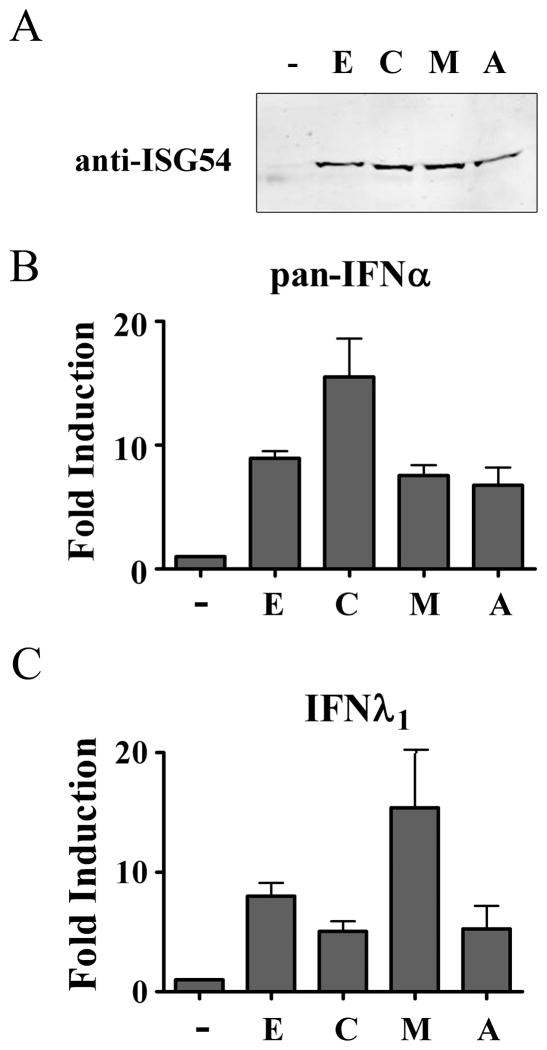
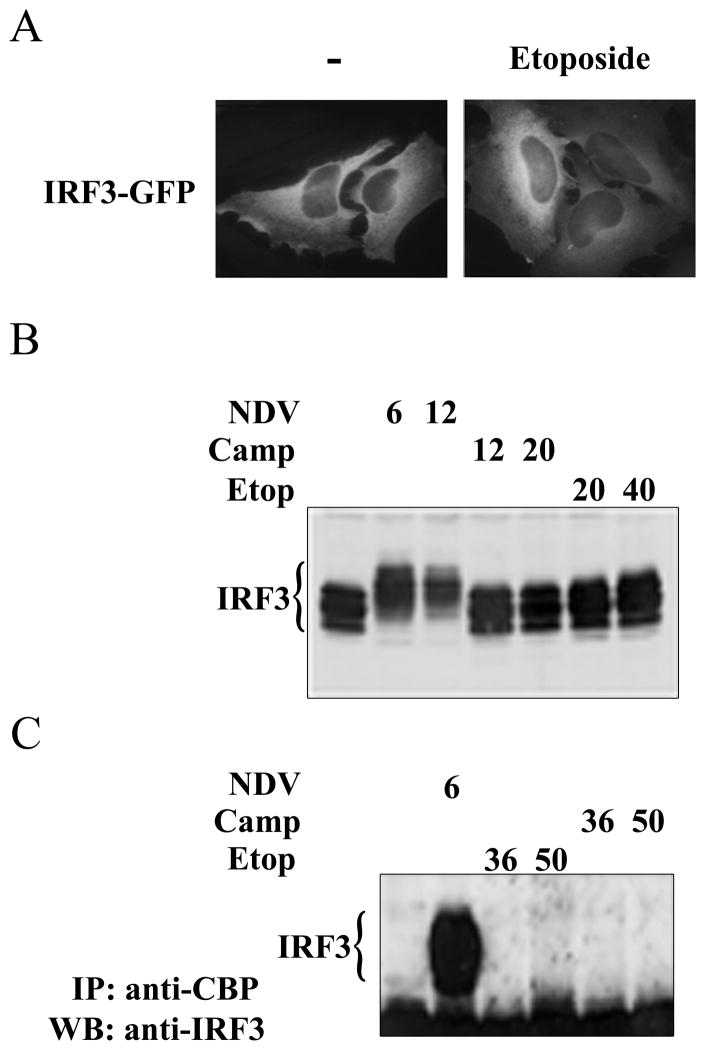
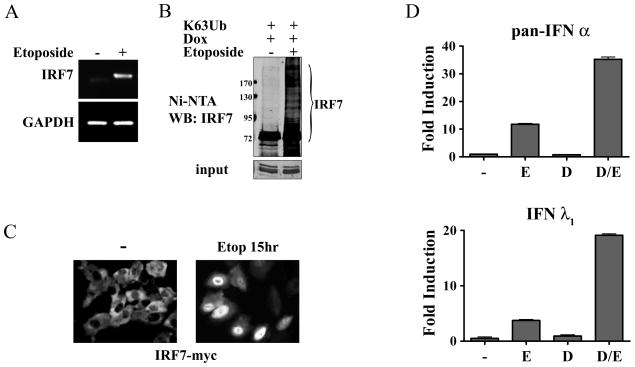
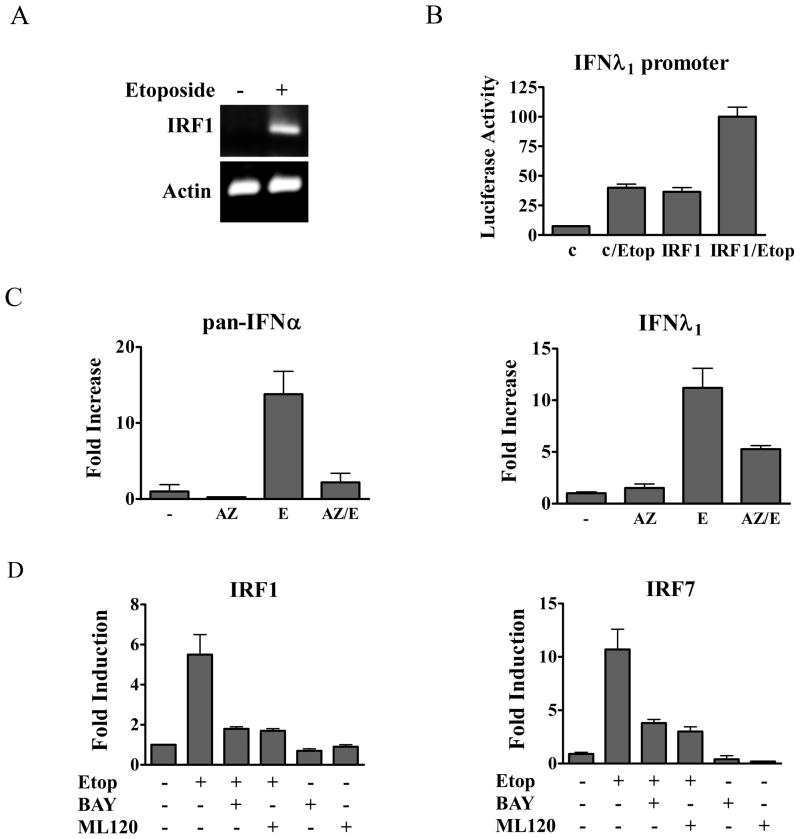
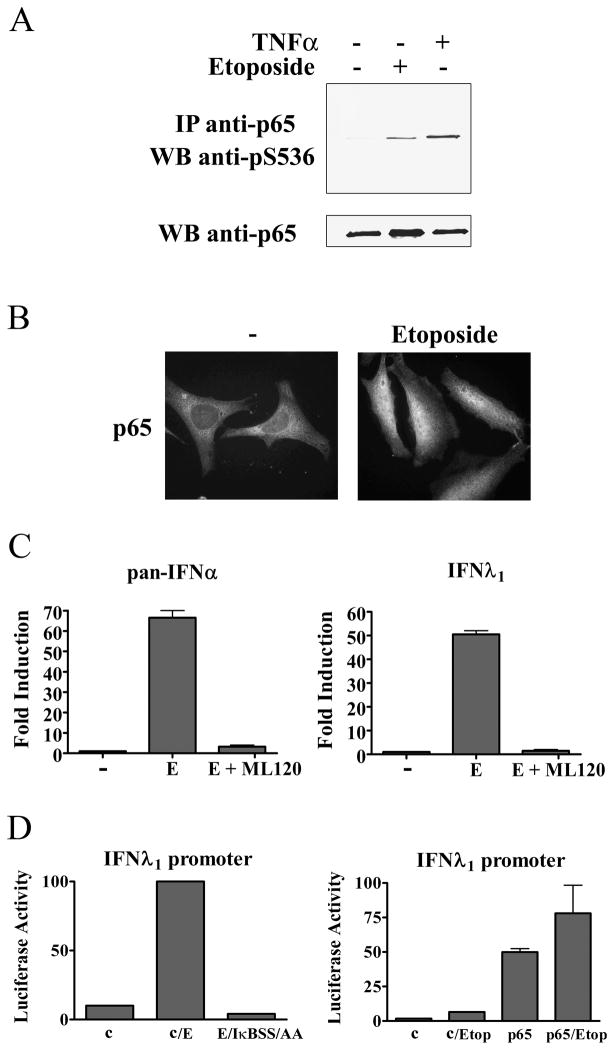
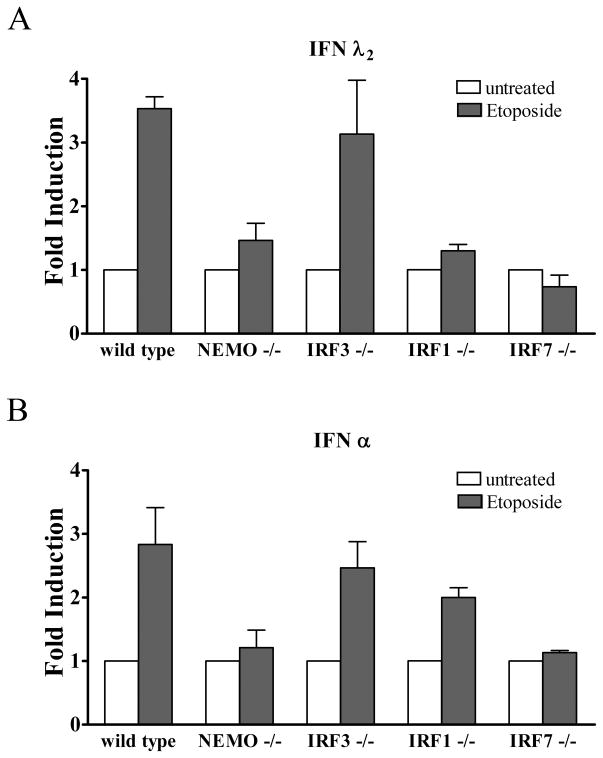
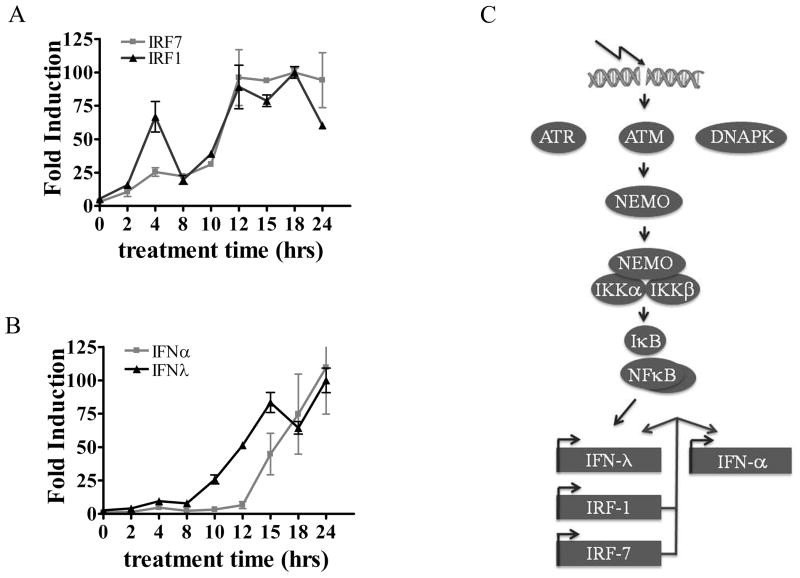
This study reveals a new complexity in the cellular response to DNA damage: activation of IFN signaling. The DNA damage response involves the rapid recruitment of repair enzymes and the activation of signal transducers that regulate cell-cycle checkpoints and cell survival. To understand the link between DNA damage and the innate cellular defense that occurs in response to many viral infections, we evaluated the effects of agents such as etoposide that promote dsDNA breaks. Treatment of human cells with etoposide led to the induction of IFN-stimulated genes and the IFN-α and IFN-λ genes. NF-κB, known to be activated in response to DNA damage, was shown to be a key regulator of this IFN gene induction. Expression of an NF-κB subunit, p65/RelA, was sufficient for induction of the human IFN-λ1 gene. In addition, NF-κB was required for the induction of IFN regulatory factor-1 and -7 that are able to stimulate expression of the IFN-α and IFN-λ genes. Cells that lack the NF-κB essential modulator lack the ability to induce the IFN genes following DNA damage. Breaks in DNA are generated during normal physiological processes of replication, transcription, and recombination, as well as by external genotoxic agents or infectious agents. The significant finding of IFN production as a stress response to DNA damage provides a new perspective on the role of IFN signaling.
Figures
References
-
- Fugmann SD, Lee AI, Shockett PE, Villey IJ, Schatz DG. The RAG proteins and V(D)J recombination: complexes, ends, and transposition. Annu Rev Immunol. 2000;18:495–527. - PubMed
-
- Richardson C, Horikoshi N, Pandita TK. The role of the DNA double-strand break response network in meiosis. DNA Repair (Amst) 2004;3:1149–1164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
