

Framing spatial cognition: neural representations of proximal and distal frames of reference and their roles in navigation
- PMID: 22013211
- PMCID: PMC3251945
- DOI: 10.1152/physrev.00021.2010
Framing spatial cognition: neural representations of proximal and distal frames of reference and their roles in navigation
Abstract
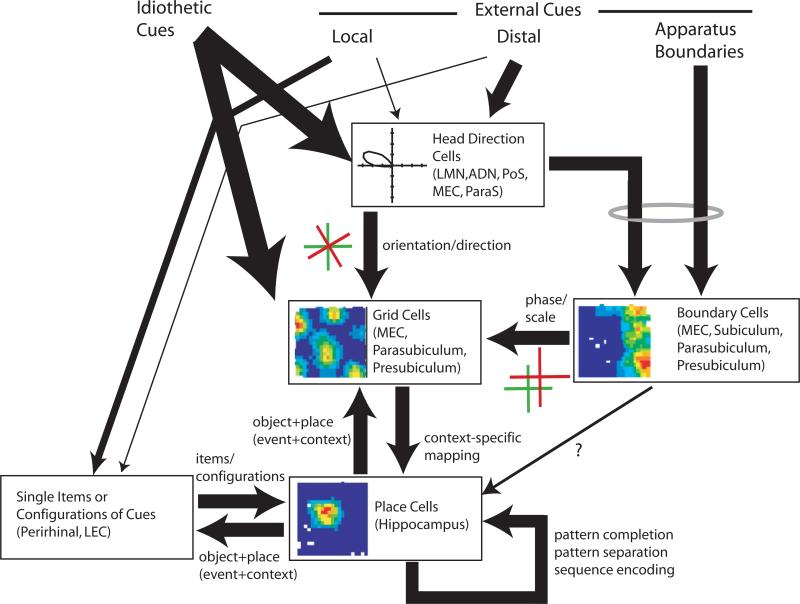
The most common behavioral test of hippocampus-dependent, spatial learning and memory is the Morris water task, and the most commonly studied behavioral correlate of hippocampal neurons is the spatial specificity of place cells. Despite decades of intensive research, it is not completely understood how animals solve the water task and how place cells generate their spatially specific firing fields. Based on early work, it has become the accepted wisdom in the general neuroscience community that distal spatial cues are the primary sources of information used by animals to solve the water task (and similar spatial tasks) and by place cells to generate their spatial specificity. More recent research, along with earlier studies that were overshadowed by the emphasis on distal cues, put this common view into question by demonstrating primary influences of local cues and local boundaries on spatial behavior and place-cell firing. This paper first reviews the historical underpinnings of the "standard" view from a behavioral perspective, and then reviews newer results demonstrating that an animal's behavior in such spatial tasks is more strongly controlled by a local-apparatus frame of reference than by distal landmarks. The paper then reviews similar findings from the literature on the neurophysiological correlates of place cells and other spatially correlated cells from related brain areas. A model is proposed by which distal cues primarily set the orientation of the animal's internal spatial coordinate system, via the head direction cell system, whereas local cues and apparatus boundaries primarily set the translation and scale of that coordinate system.
Figures
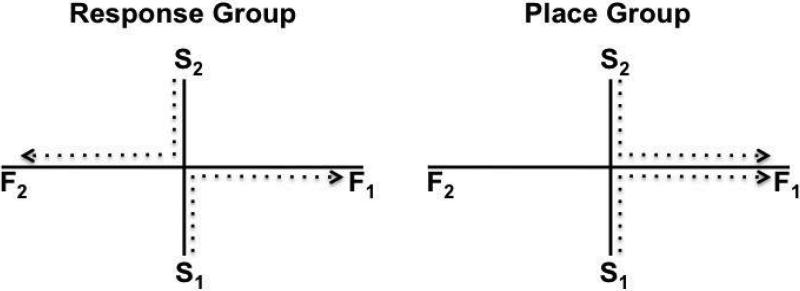
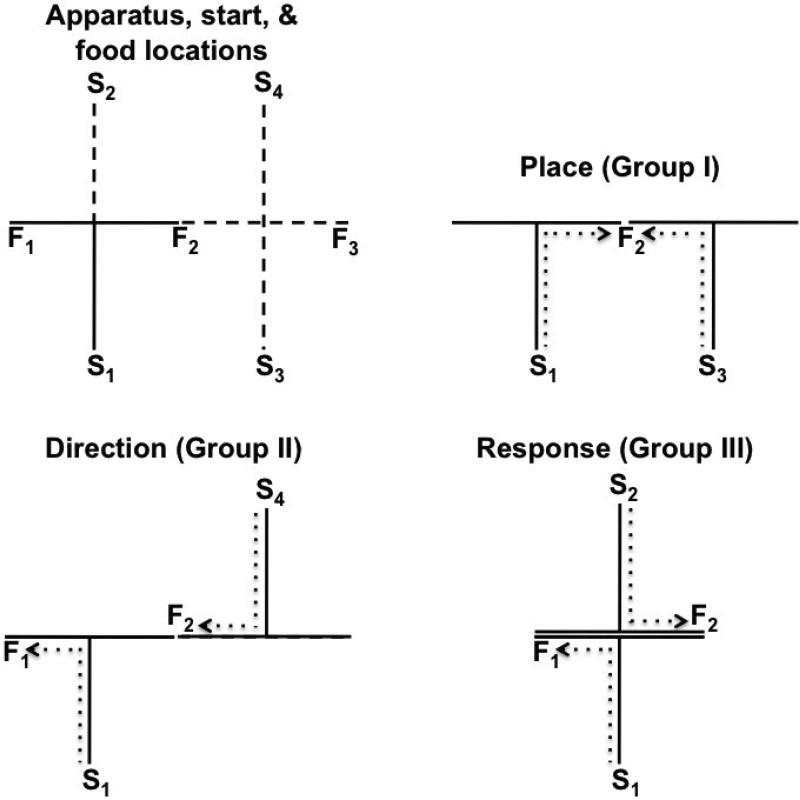
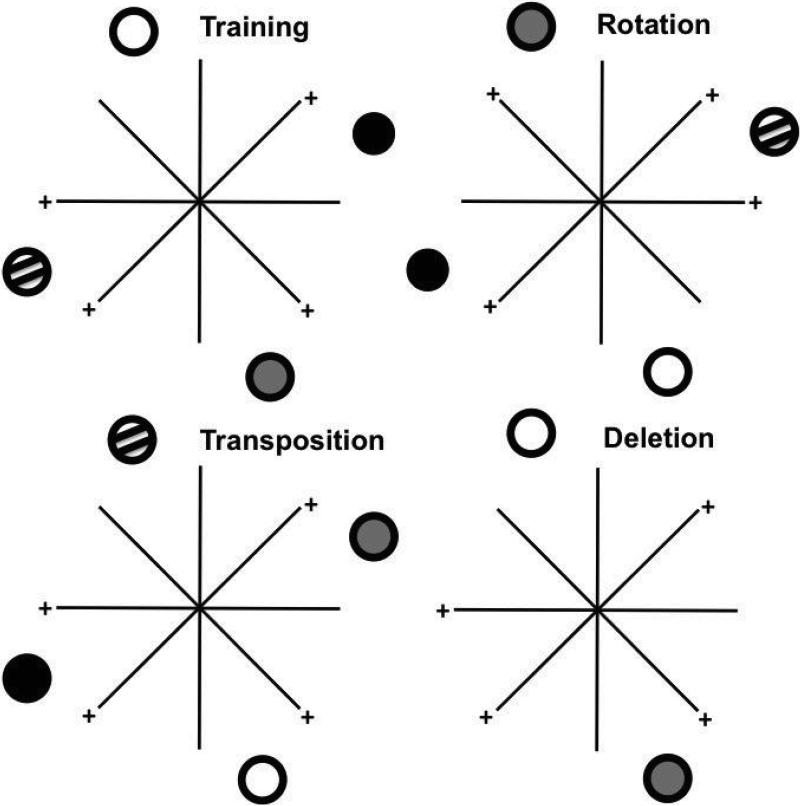
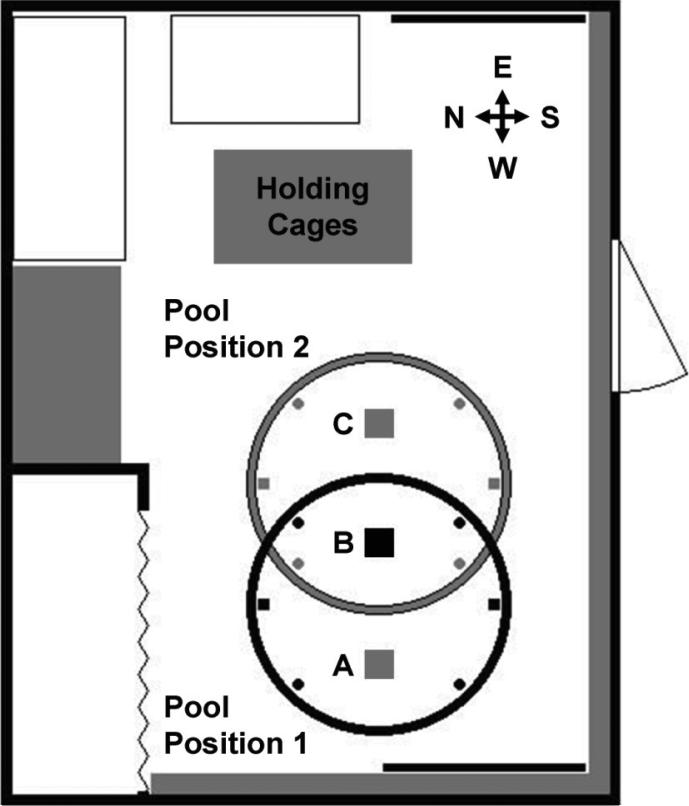
Similar articles
-
Dominance of the proximal coordinate frame in determining the locations of hippocampal place cell activity during navigation.J Neurophysiol. 2008 Jan;99(1):60-76. doi: 10.1152/jn.00731.2007. Epub 2007 Oct 24. J Neurophysiol. 2008. PMID: 17959742 Free PMC article.
-
Head direction cell representations maintain internal coherence during conflicting proximal and distal cue rotations: comparison with hippocampal place cells.J Neurosci. 2006 Jan 11;26(2):622-31. doi: 10.1523/JNEUROSCI.3885-05.2006. J Neurosci. 2006. PMID: 16407560 Free PMC article.
-
Dynamic interactions between local surface cues, distal landmarks, and intrinsic circuitry in hippocampal place cells.J Neurosci. 2002 Jul 15;22(14):6254-64. doi: 10.1523/JNEUROSCI.22-14-06254.2002. J Neurosci. 2002. PMID: 12122084 Free PMC article.
-
Place cells and place navigation.Proc Natl Acad Sci U S A. 1997 Jan 7;94(1):343-50. doi: 10.1073/pnas.94.1.343. Proc Natl Acad Sci U S A. 1997. PMID: 8990211 Free PMC article. Review.
-
Emergence of the Hippocampus as a Vector for Goal-Directed Spatial Navigation.Adv Neurobiol. 2024;41:39-61. doi: 10.1007/978-3-031-69188-1_2. Adv Neurobiol. 2024. PMID: 39589709 Review.
Cited by
-
Superficially projecting principal neurons in layer V of medial entorhinal cortex in the rat receive excitatory retrosplenial input.J Neurosci. 2013 Oct 2;33(40):15779-92. doi: 10.1523/JNEUROSCI.2646-13.2013. J Neurosci. 2013. PMID: 24089485 Free PMC article.
-
Activities of visual cortical and hippocampal neurons co-fluctuate in freely moving rats during spatial behavior.Elife. 2015 Sep 8;4:e08902. doi: 10.7554/eLife.08902. Elife. 2015. PMID: 26349031 Free PMC article.
-
Lateral Entorhinal Cortex Lesions Impair Local Spatial Frameworks.Front Syst Neurosci. 2017 May 17;11:30. doi: 10.3389/fnsys.2017.00030. eCollection 2017. Front Syst Neurosci. 2017. PMID: 28567006 Free PMC article.
-
The cognitive map in humans: spatial navigation and beyond.Nat Neurosci. 2017 Oct 26;20(11):1504-1513. doi: 10.1038/nn.4656. Nat Neurosci. 2017. PMID: 29073650 Free PMC article. Review.
-
Conflicts between local and global spatial frameworks dissociate neural representations of the lateral and medial entorhinal cortex.J Neurosci. 2013 May 29;33(22):9246-58. doi: 10.1523/JNEUROSCI.0946-13.2013. J Neurosci. 2013. PMID: 23719794 Free PMC article.
References
-
- Akers KG, Candelaria FT, Hamilton DA. Preweanling rats solve the Morris water task via directional navigation. Behavioral Neuroscience. 2007;121:1426–1430. - PubMed
-
- Akers KG, Candelaria-Cook FT, Rice JP, Johnson TE, Hamilton DA. Delayed development of place navigation compared to directional responding in young rats. Behavioral Neuroscience. 2009;123:267–275. - PubMed
-
- Arolfo MP, Nerad L, Schenk F, Bures J. Absence of snapshot memory of the target view interferes with place navigation learning by rats in the water maze. Behavioral Neuroscience. 1994;108:308–316. - PubMed
-
- Barry C, Hayman R, Burgess N, Jeffery KJ. Experience-dependent rescaling of entorhinal grids. Nat Neurosci. 2007;10:682–684. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources