

SPINDLY, a negative regulator of gibberellic acid signaling, is involved in the plant abiotic stress response
- PMID: 22013217
- PMCID: PMC3327212
- DOI: 10.1104/pp.111.187302
SPINDLY, a negative regulator of gibberellic acid signaling, is involved in the plant abiotic stress response
Abstract
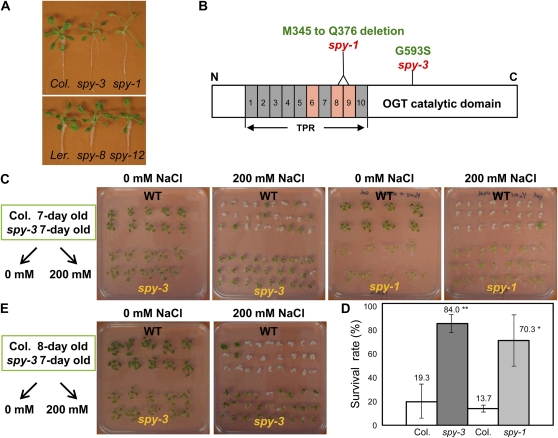
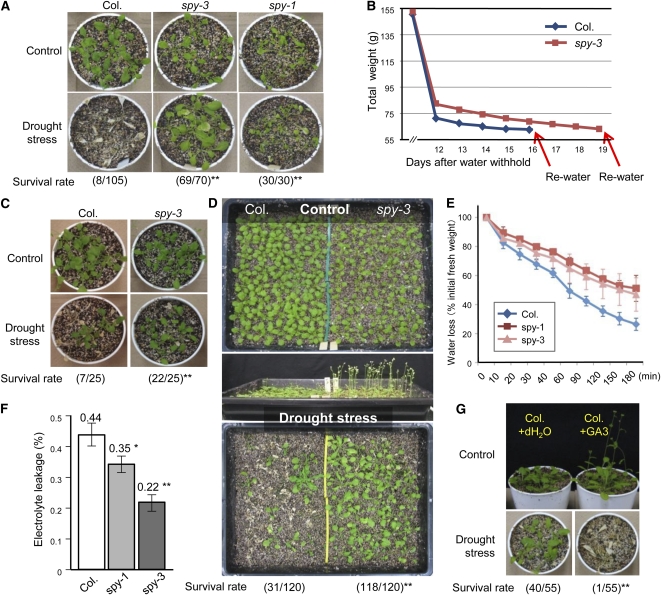
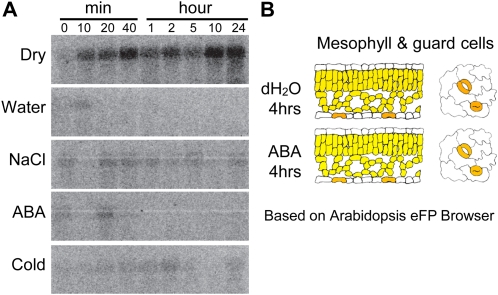
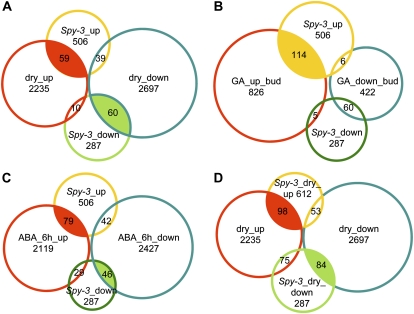
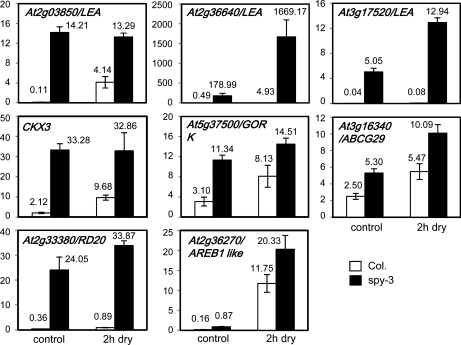
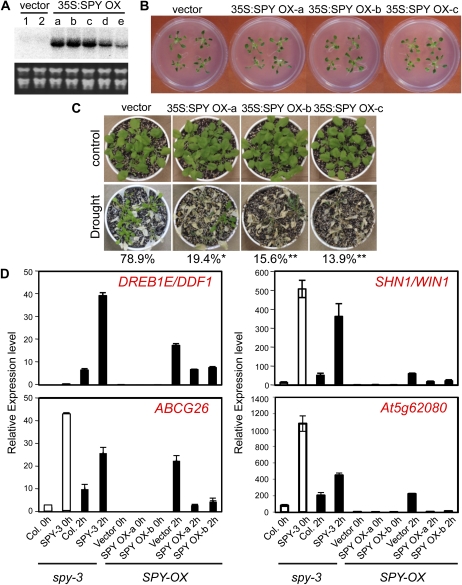
The SPINDLY (SPY) gene was first identified as a negative regulator of plant gibberellic acid (GA) signaling because mutation of this gene phenocopies plants treated with an overdose of bioactive GA and results in insensitivity to a GA inhibitor during seed germination. The SPY gene encodes an O-linked N-acetylglucosamine transferase that can modify the target protein and modulate the protein activity in cells. In this study, we describe the strong salt and drought tolerance phenotypes of Arabidopsis (Arabidopsis thaliana) spy-1 and spy-3 mutants in addition to their GA-related phenotypes. SPY gene expression was found to be drought stress inducible and slightly responsive to salt stress. Transcriptome analysis of spy-3 revealed that many GA-responsive genes were up-regulated, which could explain the GA-overdosed phenotype of spy-3. Some stress-inducible genes were found to be up-regulated in spy-3, such as genes encoding late embryogenesis abundant proteins, Responsive to Dehydration20, and AREB1-like transcription factor, which may confer stress tolerance on spy-3. CKX3, a cytokinin (CK) catabolism gene, was up-regulated in spy-3; this up-regulation indicates that the mutant possesses reduced CK signaling, which is consistent with a positive role for SPY in CK signaling. Moreover, overexpression of SPY in transgenics (SPY overexpressing [SPY-OX]) impaired plant drought stress tolerance, opposite to the phenotype of spy. The expression levels of several genes, such as DREB1E/DDF1 and SNH1/WIN1, were decreased in SPY-OX but increased in spy-3. Taken together, these data indicate that SPY plays a negative role in plant abiotic stress tolerance, probably by integrating environmental stress signals via GA and CK cross talk.
Figures
Similar articles
-
Cross talk between gibberellin and cytokinin: the Arabidopsis GA response inhibitor SPINDLY plays a positive role in cytokinin signaling.Plant Cell. 2005 Jan;17(1):92-102. doi: 10.1105/tpc.104.028472. Epub 2004 Dec 17. Plant Cell. 2005. PMID: 15608330 Free PMC article.
-
Cytosolic activity of SPINDLY implies the existence of a DELLA-independent gibberellin-response pathway.Plant J. 2009 Jun;58(6):979-88. doi: 10.1111/j.1365-313X.2009.03840.x. Epub 2009 Feb 18. Plant J. 2009. PMID: 19228341
-
The Arabidopsis gibberellin methyl transferase 1 suppresses gibberellin activity, reduces whole-plant transpiration and promotes drought tolerance in transgenic tomato.Plant Cell Environ. 2014 Jan;37(1):113-23. doi: 10.1111/pce.12135. Epub 2013 Jun 4. Plant Cell Environ. 2014. PMID: 23668385
-
Molecular and physiological mechanisms regulating tissue reunion in incised plant tissues.J Plant Res. 2015 May;128(3):381-8. doi: 10.1007/s10265-015-0705-z. Epub 2015 Mar 4. J Plant Res. 2015. PMID: 25736731 Review.
-
Gibberellic acid and cGMP-dependent transcriptional regulation in Arabidopsis thaliana.Plant Signal Behav. 2010 Mar;5(3):224-32. doi: 10.4161/psb.5.3.10718. Epub 2010 Mar 20. Plant Signal Behav. 2010. PMID: 20118660 Free PMC article. Review.
Cited by
-
Unveiling the role of epigenetic mechanisms and redox signaling in alleviating multiple abiotic stress in plants.Front Plant Sci. 2024 Sep 19;15:1456414. doi: 10.3389/fpls.2024.1456414. eCollection 2024. Front Plant Sci. 2024. PMID: 39363922 Free PMC article. Review.
-
Regulation of DELLA Proteins by Post-translational Modifications.Plant Cell Physiol. 2020 Dec 23;61(11):1891-1901. doi: 10.1093/pcp/pcaa113. Plant Cell Physiol. 2020. PMID: 32886774 Free PMC article. Review.
-
Identification and Characterization of Key Genes Responsible for Weedy and Cultivar Growth Types in Soybean.Front Genet. 2022 Feb 24;13:805347. doi: 10.3389/fgene.2022.805347. eCollection 2022. Front Genet. 2022. PMID: 35281824 Free PMC article.
-
Molecular Mechanisms of Plant Responses to Salt Stress.Front Plant Sci. 2022 Jun 27;13:934877. doi: 10.3389/fpls.2022.934877. eCollection 2022. Front Plant Sci. 2022. PMID: 35832230 Free PMC article. Review.
-
A rapid virus-induced gene silencing (VIGS) method for assessing resistance and susceptibility to cassava mosaic disease.Virol J. 2017 Mar 7;14(1):47. doi: 10.1186/s12985-017-0716-6. Virol J. 2017. PMID: 28270156 Free PMC article.
References
-
- Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP. (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94 - PubMed
-
- Comer FI, Hart GW. (2001) Reciprocity between O-GlcNAc and O-phosphate on the carboxyl terminal domain of RNA polymerase II. Biochemistry 40: 7845–7852 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
