

Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat
- PMID: 22013218
- PMCID: PMC3327217
- DOI: 10.1104/pp.111.183657
Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat
Abstract
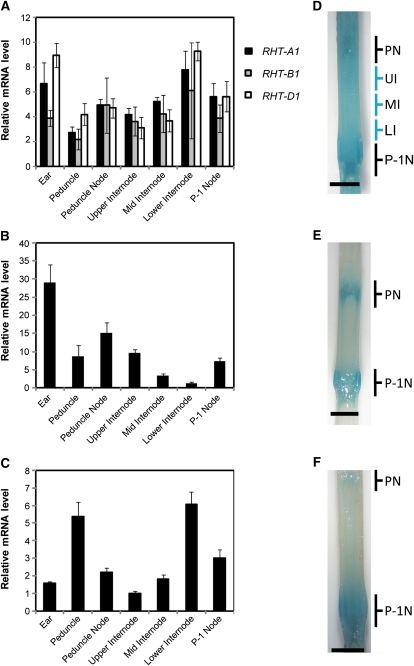
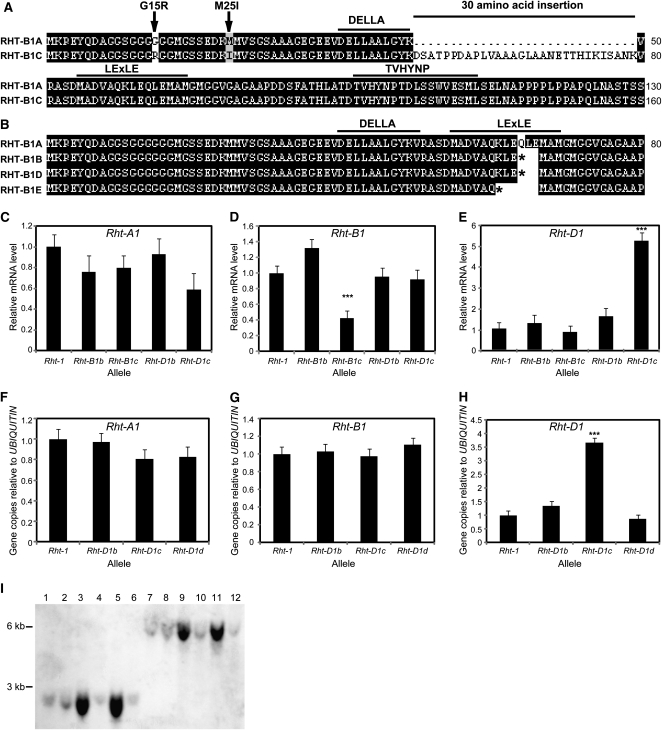
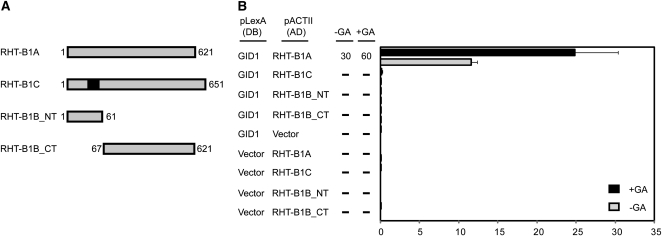
The introduction of the Reduced height (Rht)-B1b and Rht-D1b semidwarfing genes led to impressive increases in wheat (Triticum aestivum) yields during the Green Revolution. The reduction in stem elongation in varieties containing these alleles is caused by a limited response to the phytohormone gibberellin (GA), resulting in improved resistance to stem lodging and yield benefits through an increase in grain number. Rht-B1 and Rht-D1 encode DELLA proteins, which act to repress GA-responsive growth, and their mutant alleles Rht-B1b and Rht-D1b are thought to confer dwarfism by producing more active forms of these growth repressors. While no semidwarfing alleles of Rht-A1 have been identified, we show that this gene is expressed at comparable levels to the other homeologs and represents a potential target for producing novel dwarfing alleles. In this study, we have characterized additional dwarfing mutations in Rht-B1 and Rht-D1. We show that the severe dwarfism conferred by Rht-B1c is caused by an intragenic insertion, which results in an in-frame 90-bp insertion in the transcript and a predicted 30-amino acid insertion within the highly conserved amino-terminal DELLA domain. In contrast, the extreme dwarfism of Rht-D1c is due to overexpression of the semidwarfing Rht-D1b allele, caused by an increase in gene copy number. We show also that the semidwarfing alleles Rht-B1d and Rht-B1e introduce premature stop codons within the amino-terminal coding region. Yeast two-hybrid assays indicate that these newly characterized mutations in Rht-B1 and Rht-D1 confer "GA-insensitive" dwarfism by producing DELLA proteins that do not bind the GA receptor GA INSENSITIVE DWARF1, potentially compromising their targeted degradation.
Figures


References
-
- Appleford NEJ, Evans DJ, Lenton JR, Gaskin P, Croker SJ, Devos KM, Phillips AL, Hedden P. (2006) Function and transcript analysis of gibberellin-biosynthetic enzymes in wheat. Planta 223: 568–582 - PubMed
-
- Asano K, Hirano K, Ueguchi-Tanaka M, Angeles-Shim RB, Komura T, Satoh H, Kitano H, Matsuoka M, Ashikari M. (2009) Isolation and characterization of dominant dwarf mutants, Slr1-d, in rice. Mol Genet Genomics 281: 223–231 - PubMed
-
- Borner A, Lehmann CO, Mettin J, Plaschke J, Schlegel G, Schlegel R, Melz G, Thiele V. (1991) GA-insensitivity of ‘Aibian 1a’: pleiotropic effects of isogenic Rht-lines. Ann Wheat Newsletter 37: 59–60
-
- Borner A, Plaschke J, Korzun V, Worland AJ. (1996) The relationships between the dwarfing genes of wheat and rye. Euphytica 89: 69–75
-
- Boss PK, Thomas MR. (2002) Association of dwarfism and floral induction with a grape ‘Green Revolution’ mutation. Nature 416: 847–850 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- BB/E006922/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/J/00000606/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E00704X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/C/00004951/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/D019001/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
