

S(+)amphetamine induces a persistent leak in the human dopamine transporter: molecular stent hypothesis
- PMID: 22014068
- PMCID: PMC3423253
- DOI: 10.1111/j.1476-5381.2011.01728.x
S(+)amphetamine induces a persistent leak in the human dopamine transporter: molecular stent hypothesis
Abstract
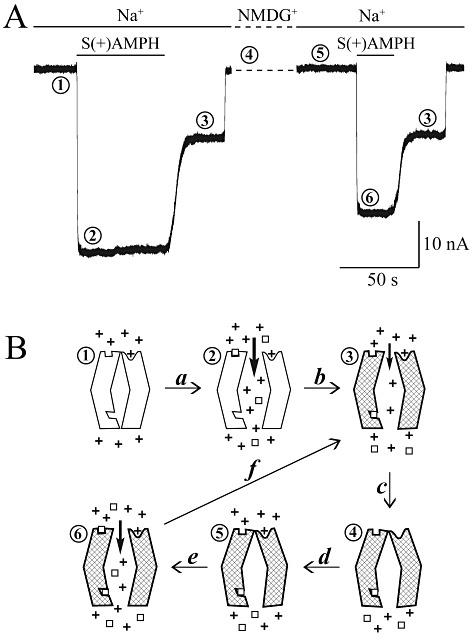
Background and purpose: Wherever they are located, dopamine transporters (DATs) clear dopamine (DA) from the extracellular milieu to help regulate dopaminergic signalling. Exposure to amphetamine (AMPH) increases extracellular DA in the synaptic cleft, which has been ascribed to DAT reverse transport. Increased extracellular DA prolongs postsynaptic activity and reinforces abuse and hedonic behaviour.
Experimental approach: Xenopus laevis oocytes expressing human (h) DAT were voltage-clamped and exposed to DA, R(-)AMPH, or S(+)AMPH.
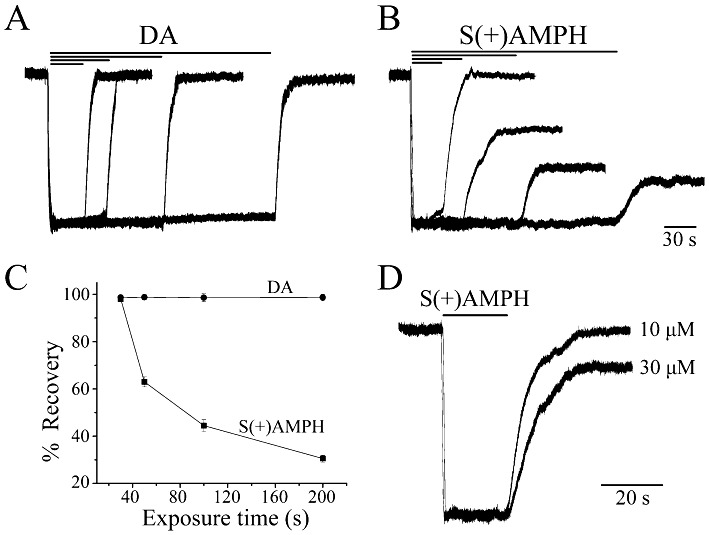
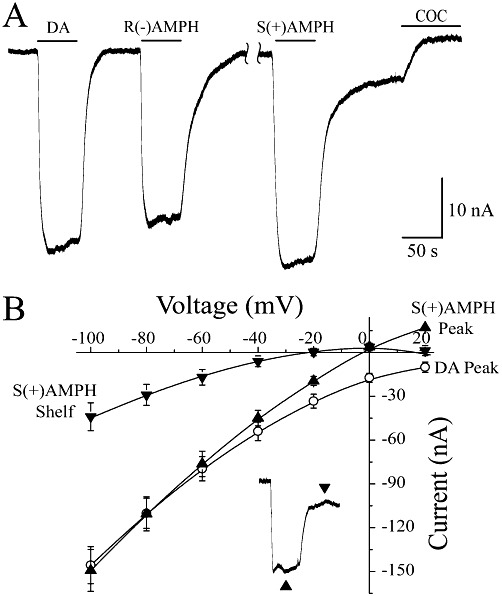
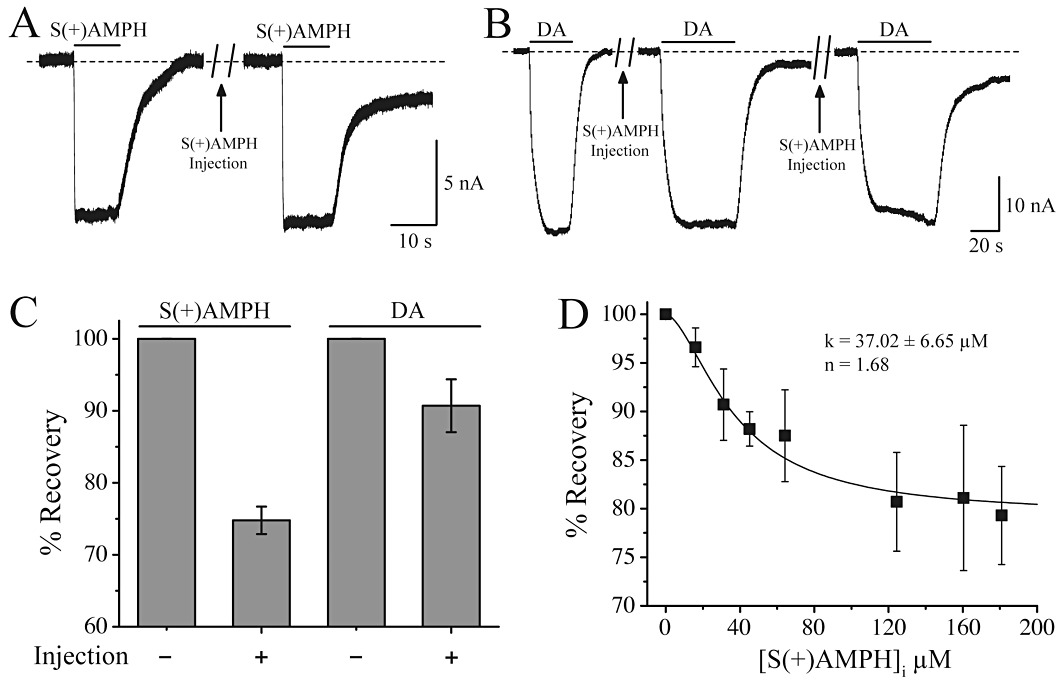
Key results: At -60mV, near neuronal resting potentials, S(+)AMPH induced a depolarizing current through hDAT, which after removing the drug, persisted for more than 30 min. This persistent leak in the absence of S(+)AMPH was in contrast to the currents induced by R(-)AMPH and DA, which returned to baseline immediately after their removal. Our data suggest that S(+)AMPH and Na(+) carry the initial S(+)AMPH-induced current, whereas Na+ and Cl(-) carry the persistent leak current. We propose that the persistent current results from the internal action of S(+)AMPH on hDAT because the temporal effect was consistent with S(+)AMPH influx, and intracellular S(+)AMPH activated the effect. The persistent current was dependent on Na(+) and was blocked by cocaine. Intracellular injection of S(+)AMPH also activated a DA-induced persistent leak current.
Conclusions and implications: We report a hitherto unknown action of S(+)AMPH on hDAT that potentially affects AMPH-induced DA release. We propose that internal S(+)AMPH acts as a molecular stent that holds the transporter open even after external S(+)AMPH is removed. Amphetamine-induced persistent leak currents are likely to influence dopaminergic signalling, DA release mechanisms, and amphetamine abuse.
© 2011 The Authors. British Journal of Pharmacology © 2011 The British Pharmacological Society.
Figures
References
-
- Amara SG, Sonders MS. Neurotransmitter transporters as molecular targets for addictive drugs. Drug Alcohol Depend. 1998;51:87–96. - PubMed
-
- Burnette WB, Bailey MD, Kukoyi S, Blakely RD, Trowbridge CG, Justice JB., Jr Human norepinephrine transporter kinetics using rotating disk electrode voltammetry. Anal Chem. 1996;68:2932–2938. - PubMed
-
- De Felice LJ. Evaluation of: Piscitelli CL et al. Nature 2010. 2011;468:1129–1132. F1000.com/7439956. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
