

The complexity of cardiolipin in health and disease
- PMID: 22014644
- PMCID: PMC3259273
- DOI: 10.1016/j.tibs.2011.09.003
The complexity of cardiolipin in health and disease
Abstract
Cardiolipin, the signature phospholipid of mitochondria, is a lipid dimer that is important for a diverse range of mitochondrial activities beyond the process of ATP production. Thus not surprisingly, derangements in cardiolipin metabolism are now appreciated to contribute to an assortment of pathological conditions. A comprehensive inventory of enzymes involved in cardiolipin biosynthesis and remodeling was just recently obtained. Post-biosynthesis, the acyl chain composition of cardiolipin is modified by up to three distinct remodeling enzymes that produce either a homogeneous tissue-specific mature form of cardiolipin or alternatively 'bad' cardiolipin that has been linked to mitochondrial dysfunction. In this review, we initially focus on the newly identified players in cardiolipin metabolism and then shift our attention to how changes in cardiolipin metabolism contribute to human disease.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
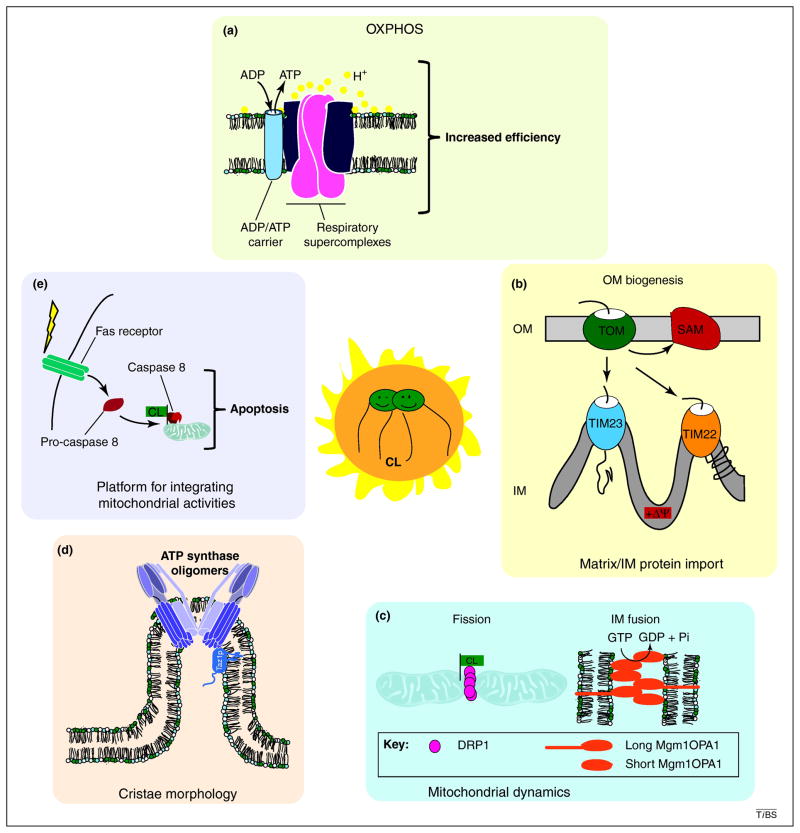
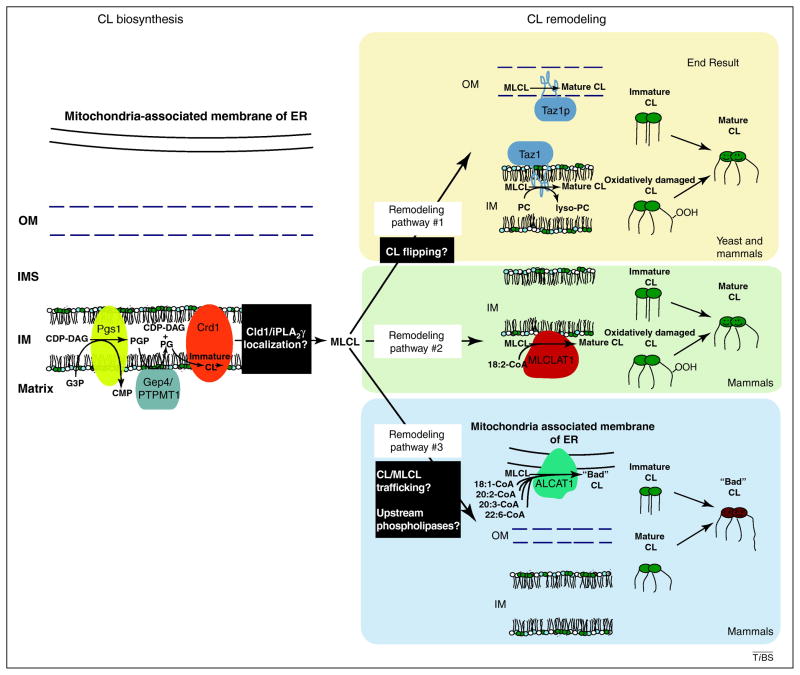
References
-
- Sparagna GC, Lesnefsky EJ. Cardiolipin remodeling in the heart. J Cardiovasc Pharmacol. 2009;53:290–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
