

The transcription factor SP4 is reduced in postmortem cerebellum of bipolar disorder subjects: control by depolarization and lithium
- PMID: 22017217
- PMCID: PMC3202296
- DOI: 10.1111/j.1399-5618.2011.00941.x
The transcription factor SP4 is reduced in postmortem cerebellum of bipolar disorder subjects: control by depolarization and lithium
Abstract
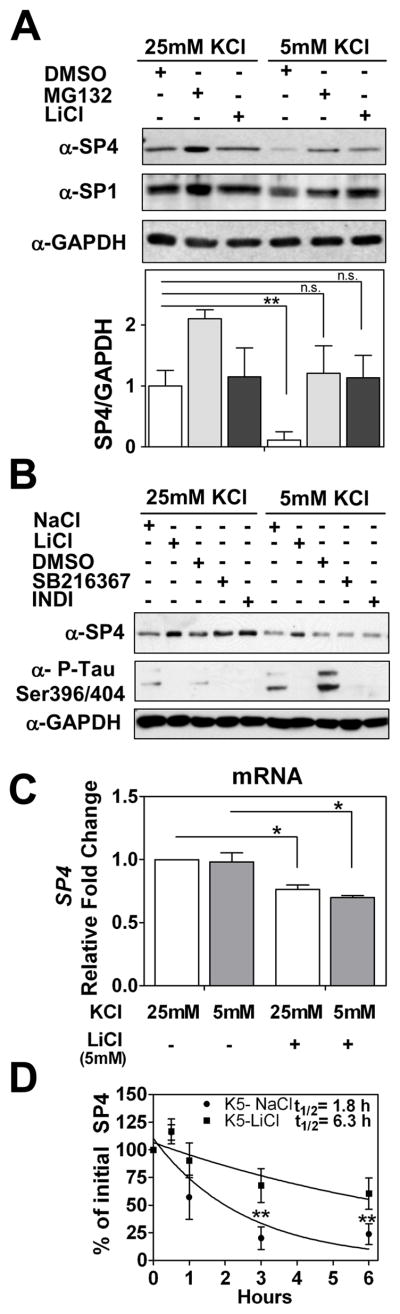
Objectives: Regulation of gene expression is important for the development and function of the nervous system. However, the transcriptional programs altered in psychiatric diseases are not completely characterized. Human gene association studies and analysis of mutant mice suggest that the transcription factor specificity protein 4 (SP4) may be implicated in the pathophysiology of psychiatric diseases. We hypothesized that SP4 levels may be altered in the brain of bipolar disorder (BD) subjects and regulated by neuronal activity and drug treatment.
Methods: We analyzed messenger RNA (mRNA) and protein levels of SP4 and SP1 in the postmortem prefrontal cortex and cerebellum of BD subjects (n = 10) and controls (n = 10). We also examined regulation of SP4 mRNA and protein levels by neuronal activity and lithium in rat cerebellar granule neurons.
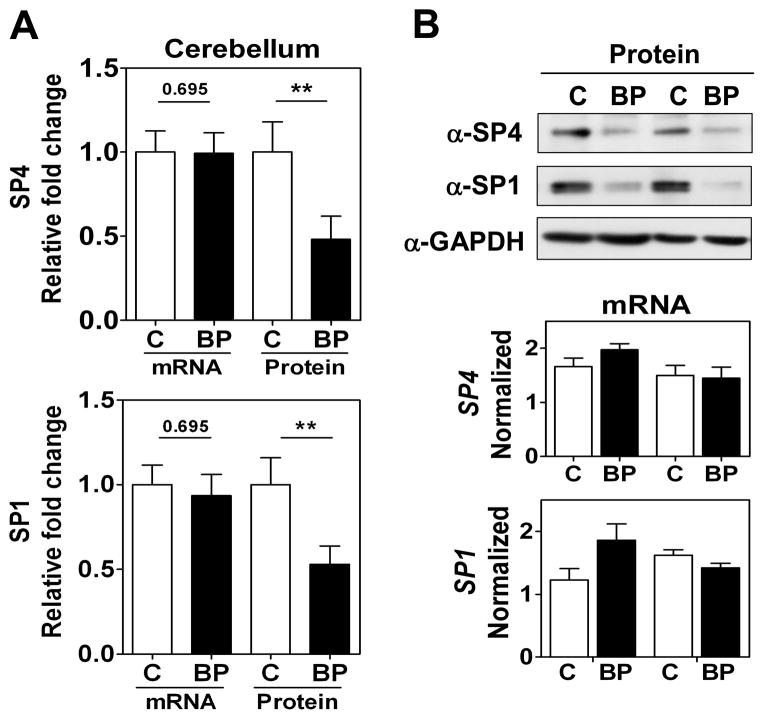
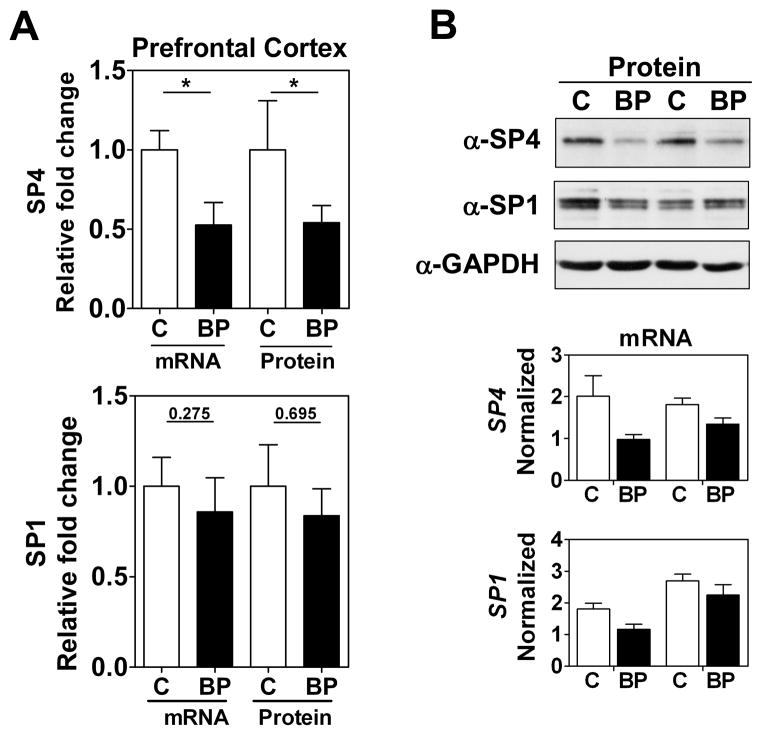
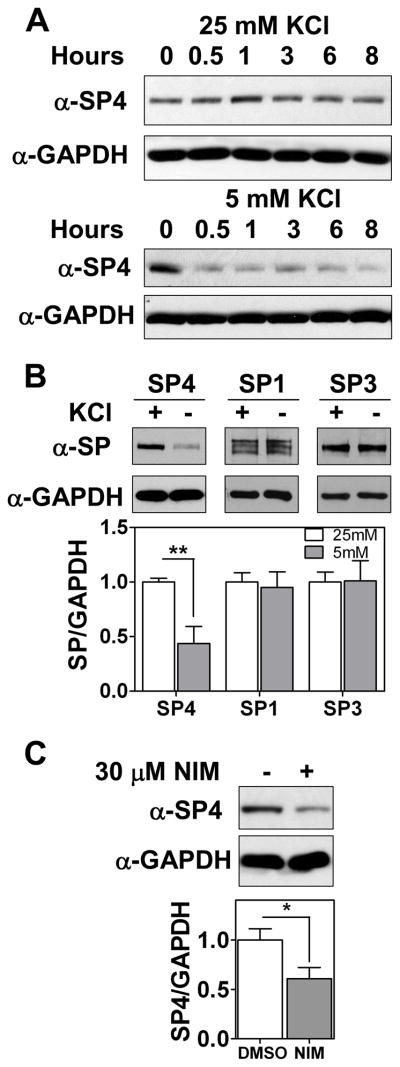
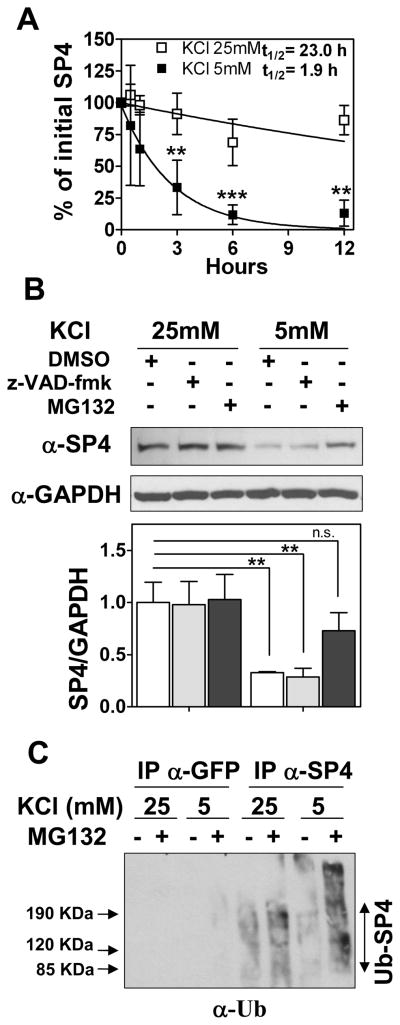
Results: We report a reduction of SP4 and SP1 proteins, but not mRNA levels, in the cerebellum of BD subjects. SP4 protein and mRNA levels were also reduced in the prefrontal cortex. Moreover, we found in rat cerebellar granule neurons that under non-depolarizing conditions SP4, but not SP1, was polyubiquitinated and degraded by the proteasome while lithium stabilized SP4 protein.
Conclusions: Our study provides the first evidence of altered SP4 protein in the cerebellum and prefrontal cortex in BD subjects supporting a possible role of transcription factor SP4 in the pathogenesis of the disease. In addition, our finding that SP4 stability is regulated by depolarization and lithium provides a pathway through which neuronal activity and lithium could control gene expression suggesting that normalization of SP4 levels could contribute to treatment of affective disorders.
© 2011 John Wiley and Sons A/S.
Conflict of interest statement
The authors of this paper do not have any commercial associations that might pose a conflict of interest in connection with this manuscript.
Figures
References
-
- Haldane M, Frangou S. New insights help define the pathophysiology of bipolar affective disorder: neuroimaging and neuropathology findings. Prog Neuropsychopharmacol Biol Psychiatry. 2004;28:943–960. - PubMed
-
- Hajek T, Carrey N, Alda M. Neuroanatomical abnormalities as risk factors for bipolar disorder. Bipolar Disord. 2005;7:393–403. - PubMed
-
- Beyer JL, Krishnan KR. Volumetric brain imaging findings in mood disorders. Bipolar Disord. 2002;4:89–104. - PubMed
-
- Bolbecker AR, Mehta C, Johannesen JK, et al. Eyeblink conditioning anomalies in bipolar disorder suggest cerebellar dysfunction. Bipolar Disord. 2009;11:19–32. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
