

Substrate recognition by β-ketoacyl-ACP synthases
- PMID: 22017312
- PMCID: PMC3361727
- DOI: 10.1021/bi201199x
Substrate recognition by β-ketoacyl-ACP synthases
Abstract
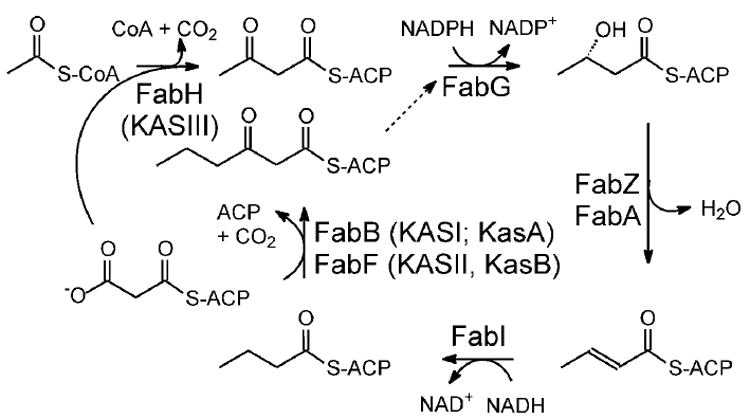
β-Ketoacyl-ACP synthase (KAS) enzymes catalyze Claisen condensation reactions in the fatty acid biosynthesis pathway. These reactions follow a ping-pong mechanism in which a donor substrate acylates the active site cysteine residue after which the acyl group is condensed with the malonyl-ACP acceptor substrate to form a β-ketoacyl-ACP. In the priming KASIII enzymes the donor substrate is an acyl-CoA while in the elongating KASI and KASII enzymes the donor is an acyl-ACP. Although the KASIII enzyme in Escherichia coli (ecFabH) is essential, the corresponding enzyme in Mycobacterium tuberculosis (mtFabH) is not, suggesting that the KASI or II enzyme in M. tuberculosis (KasA or KasB, respectively) must be able to accept a CoA donor substrate. Since KasA is essential, the substrate specificity of this KASI enzyme has been explored using substrates based on phosphopantetheine, CoA, ACP, and AcpM peptide mimics. This analysis has been extended to the KASI and KASII enzymes from E. coli (ecFabB and ecFabF) where we show that a 14-residue malonyl-phosphopantetheine peptide can efficiently replace malonyl-ecACP as the acceptor substrate in the ecFabF reaction. While ecFabF is able to catalyze the condensation reaction when CoA is the carrier for both substrates, the KASI enzymes ecFabB and KasA have an absolute requirement for an ACP substrate as the acyl donor. Provided that this requirement is met, variation in the acceptor carrier substrate has little impact on the k(cat)/K(m) for the KASI reaction. For the KASI enzymes we propose that the binding of ecACP (AcpM) results in a conformational change that leads to an open form of the enzyme to which the malonyl acceptor substrate binds. Finally, the substrate inhibition observed when palmitoyl-CoA is the donor substrate for the KasA reaction has implications for the importance of mtFabH in the mycobacterial FASII pathway.
Figures
Similar articles
-
Purification and biochemical characterization of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthases KasA and KasB.J Biol Chem. 2001 Dec 14;276(50):47029-37. doi: 10.1074/jbc.M108903200. Epub 2001 Oct 12. J Biol Chem. 2001. PMID: 11600501
-
Slow onset inhibition of bacterial beta-ketoacyl-acyl carrier protein synthases by thiolactomycin.J Biol Chem. 2010 Feb 26;285(9):6161-9. doi: 10.1074/jbc.M109.077909. Epub 2009 Dec 16. J Biol Chem. 2010. PMID: 20018879 Free PMC article.
-
The Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III activity is inhibited by phosphorylation on a single threonine residue.J Biol Chem. 2009 Mar 6;284(10):6414-24. doi: 10.1074/jbc.M806537200. Epub 2008 Dec 11. J Biol Chem. 2009. PMID: 19074144 Free PMC article.
-
Structural classification and properties of ketoacyl synthases.Protein Sci. 2011 Oct;20(10):1659-67. doi: 10.1002/pro.712. Protein Sci. 2011. PMID: 21830247 Free PMC article. Review.
-
The β-ketoacyl-ACP synthase from Mycobacterium tuberculosis as potential drug targets.Curr Med Chem. 2011;18(9):1318-24. doi: 10.2174/092986711795029636. Curr Med Chem. 2011. PMID: 21370994 Review.
Cited by
-
Metabolic rearrangement enables adaptation of microbial growth rate to temperature shifts.Nat Microbiol. 2025 Jan;10(1):185-201. doi: 10.1038/s41564-024-01841-4. Epub 2024 Dec 13. Nat Microbiol. 2025. PMID: 39672961
-
A kinetic framework for modeling oleochemical biosynthesis in Escherichia coli.Biotechnol Bioeng. 2022 Nov;119(11):3149-3161. doi: 10.1002/bit.28209. Epub 2022 Aug 24. Biotechnol Bioeng. 2022. PMID: 35959746 Free PMC article.
-
Structural basis for the recognition of mycolic acid precursors by KasA, a condensing enzyme and drug target from Mycobacterium tuberculosis.J Biol Chem. 2013 Nov 22;288(47):34190-34204. doi: 10.1074/jbc.M113.511436. Epub 2013 Oct 9. J Biol Chem. 2013. PMID: 24108128 Free PMC article.
-
A Canonical Biotin Synthesis Enzyme, 8-Amino-7-Oxononanoate Synthase (BioF), Utilizes Different Acyl Chain Donors in Bacillus subtilis and Escherichia coli.Appl Environ Microbiol. 2017 Dec 15;84(1):e02084-17. doi: 10.1128/AEM.02084-17. Print 2018 Jan 1. Appl Environ Microbiol. 2017. PMID: 29054876 Free PMC article.
-
A Preclinical Candidate Targeting Mycobacterium tuberculosis KasA.Cell Chem Biol. 2020 May 21;27(5):560-570.e10. doi: 10.1016/j.chembiol.2020.02.007. Epub 2020 Mar 19. Cell Chem Biol. 2020. PMID: 32197094 Free PMC article.
References
-
- McDevitt D, Rosenberg M. Exploiting genomics to discover new antibiotics. Trends Microbiol. 2001;9:611–617. - PubMed
-
- Payne DJ, Warren PV, Holmes DJ, Ji Y, Lonsdale JT. Bacterial fatty-acid biosynthesis: a genomics-driven target for antibacterial drug discovery. Drug Discov Today. 2001;6:537–544. - PubMed
-
- Campbell JW, Cronan JE., Jr Bacterial fatty acid biosynthesis: targets for antibacterial drug discovery. Annu Rev Microbiol. 2001;55:305–332. - PubMed
-
- Banerjee A, Dubnau E, Quemard A, Balasubramanian V, Um KS, Wilson T, Collins D, de Lisle G, Jacobs WR., Jr inhA, a gene encoding a target for isoniazid and ethionamide in Mycobacterium tuberculosis. Science. 1994;263:227–230. - PubMed
-
- Dessen A, Quemard A, Blanchard JS, Jacobs WR, Jr, Sacchettini JC. Crystal structure and function of the isoniazid target of Mycobacterium tuberculosis. Science. 1995;267:1638–1641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
