

Substrate recognition by β-ketoacyl-ACP synthases
- PMID: 22017312
- PMCID: PMC3361727
- DOI: 10.1021/bi201199x
Substrate recognition by β-ketoacyl-ACP synthases
Abstract
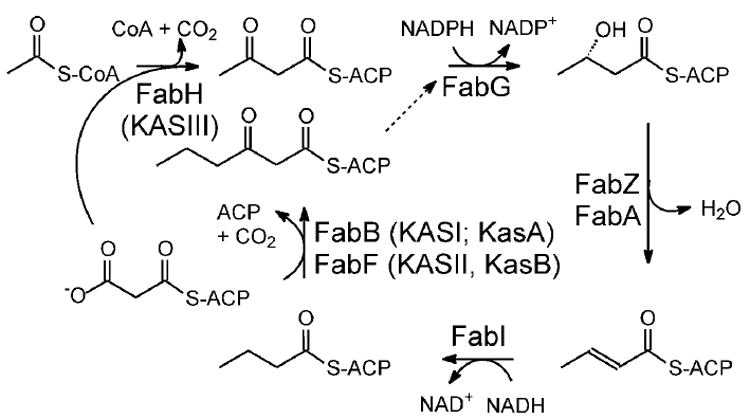
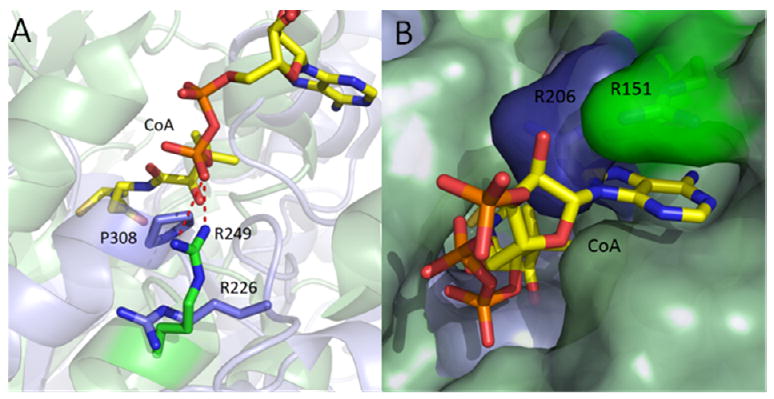
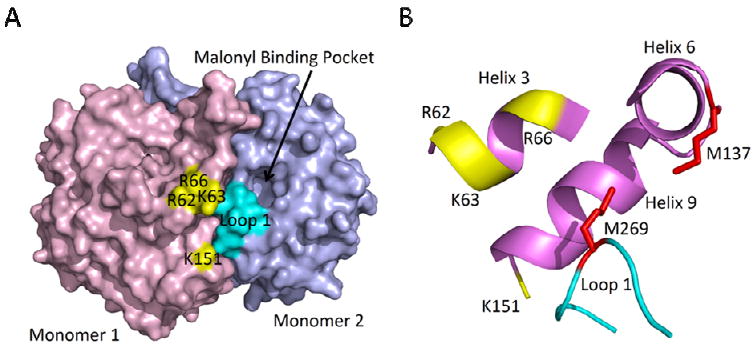
β-Ketoacyl-ACP synthase (KAS) enzymes catalyze Claisen condensation reactions in the fatty acid biosynthesis pathway. These reactions follow a ping-pong mechanism in which a donor substrate acylates the active site cysteine residue after which the acyl group is condensed with the malonyl-ACP acceptor substrate to form a β-ketoacyl-ACP. In the priming KASIII enzymes the donor substrate is an acyl-CoA while in the elongating KASI and KASII enzymes the donor is an acyl-ACP. Although the KASIII enzyme in Escherichia coli (ecFabH) is essential, the corresponding enzyme in Mycobacterium tuberculosis (mtFabH) is not, suggesting that the KASI or II enzyme in M. tuberculosis (KasA or KasB, respectively) must be able to accept a CoA donor substrate. Since KasA is essential, the substrate specificity of this KASI enzyme has been explored using substrates based on phosphopantetheine, CoA, ACP, and AcpM peptide mimics. This analysis has been extended to the KASI and KASII enzymes from E. coli (ecFabB and ecFabF) where we show that a 14-residue malonyl-phosphopantetheine peptide can efficiently replace malonyl-ecACP as the acceptor substrate in the ecFabF reaction. While ecFabF is able to catalyze the condensation reaction when CoA is the carrier for both substrates, the KASI enzymes ecFabB and KasA have an absolute requirement for an ACP substrate as the acyl donor. Provided that this requirement is met, variation in the acceptor carrier substrate has little impact on the k(cat)/K(m) for the KASI reaction. For the KASI enzymes we propose that the binding of ecACP (AcpM) results in a conformational change that leads to an open form of the enzyme to which the malonyl acceptor substrate binds. Finally, the substrate inhibition observed when palmitoyl-CoA is the donor substrate for the KasA reaction has implications for the importance of mtFabH in the mycobacterial FASII pathway.
Figures
References
-
- McDevitt D, Rosenberg M. Exploiting genomics to discover new antibiotics. Trends Microbiol. 2001;9:611–617. - PubMed
-
- Payne DJ, Warren PV, Holmes DJ, Ji Y, Lonsdale JT. Bacterial fatty-acid biosynthesis: a genomics-driven target for antibacterial drug discovery. Drug Discov Today. 2001;6:537–544. - PubMed
-
- Campbell JW, Cronan JE., Jr Bacterial fatty acid biosynthesis: targets for antibacterial drug discovery. Annu Rev Microbiol. 2001;55:305–332. - PubMed
-
- Banerjee A, Dubnau E, Quemard A, Balasubramanian V, Um KS, Wilson T, Collins D, de Lisle G, Jacobs WR., Jr inhA, a gene encoding a target for isoniazid and ethionamide in Mycobacterium tuberculosis. Science. 1994;263:227–230. - PubMed
-
- Dessen A, Quemard A, Blanchard JS, Jacobs WR, Jr, Sacchettini JC. Crystal structure and function of the isoniazid target of Mycobacterium tuberculosis. Science. 1995;267:1638–1641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
