

Review
doi: 10.1021/cr200073d.
Epub 2011 Oct 21.
Enzymatic chemistry of cyclopropane, epoxide, and aziridine biosynthesis
Affiliations
- PMID: 22017381
- PMCID: PMC3288687
- DOI: 10.1021/cr200073d
Item in Clipboard
Review
Enzymatic chemistry of cyclopropane, epoxide, and aziridine biosynthesis
Chem Rev.
.
No abstract available
Figures
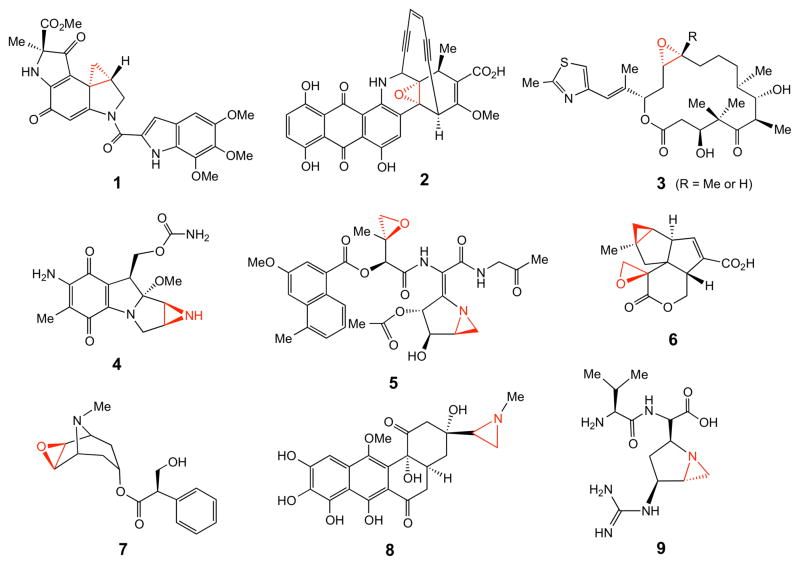
Representative three membered ring-containing natural products.
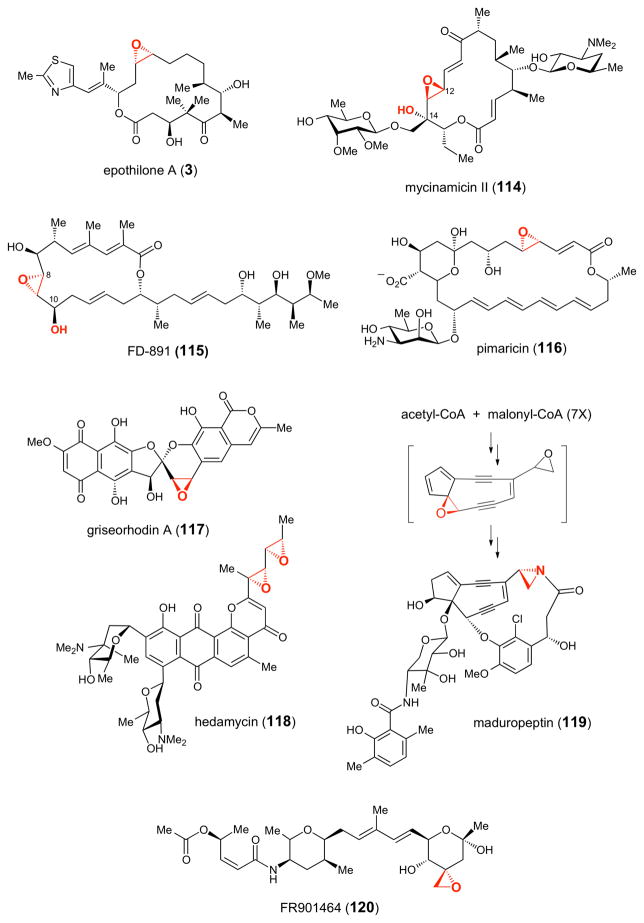
Representative epoxide-containing natural products whose biosynthetic gene clusters contain genes encoding putative P450 epoxidases.
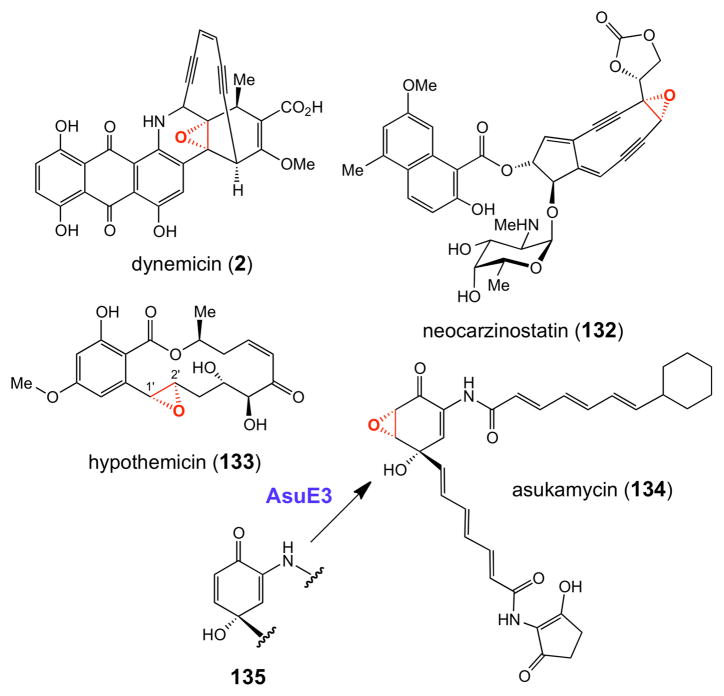
Representative natural products whose epoxide moieties are likely installed by flavindependent epoxidases. See also hedamycin (118) in Figure 2.
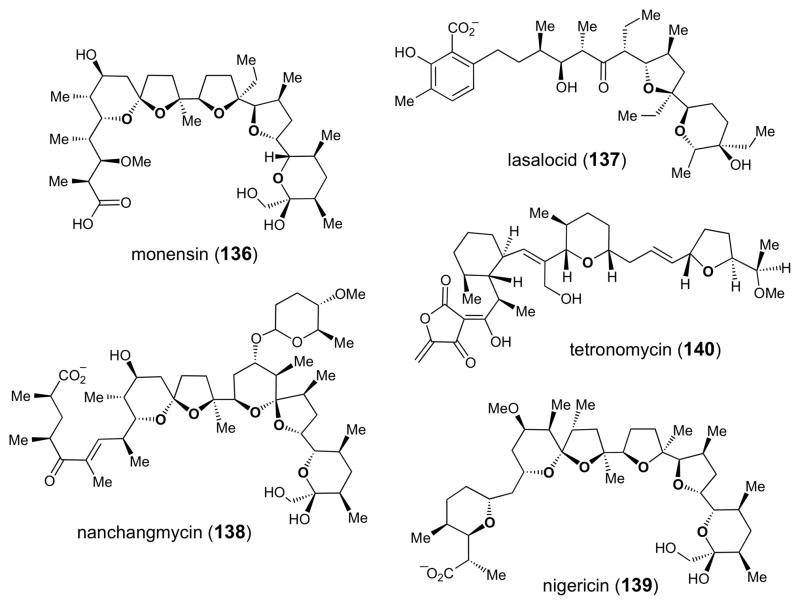
Chemical structures of several polyether natural products whose biosynthetic gene clusters have been sequenced.
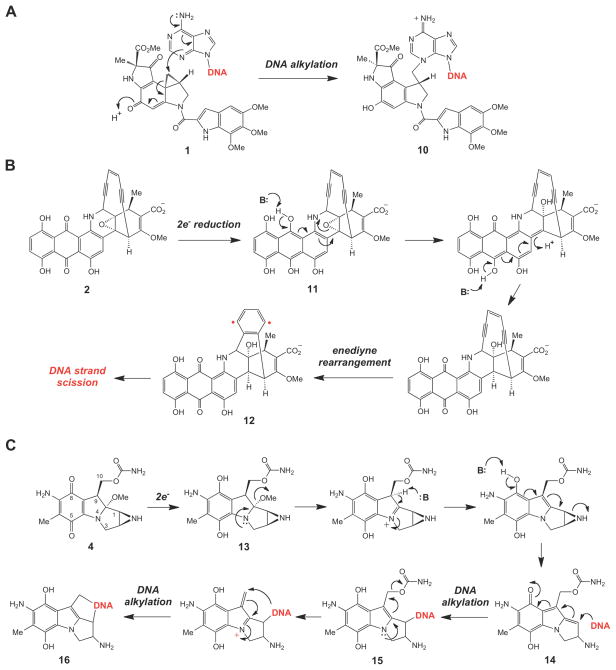
Mechanism of action of (A) duocarmycin, (B) dynemicin, and (C) mitomycin.
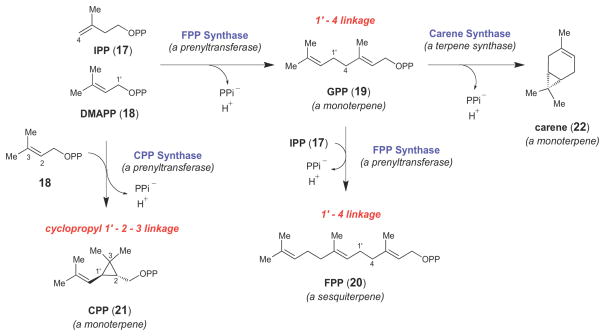
Representative examples of prenyltransferase- and terpene synthase-catalyzed reactions.
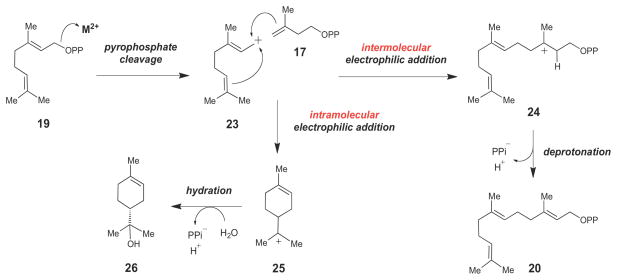
Mechanistic link between prenyltransferases and terpene synthases. Both families of enzymes catalyze C-C bond formation via reactive carbocation intermediates (such as 23, 24 and 25).
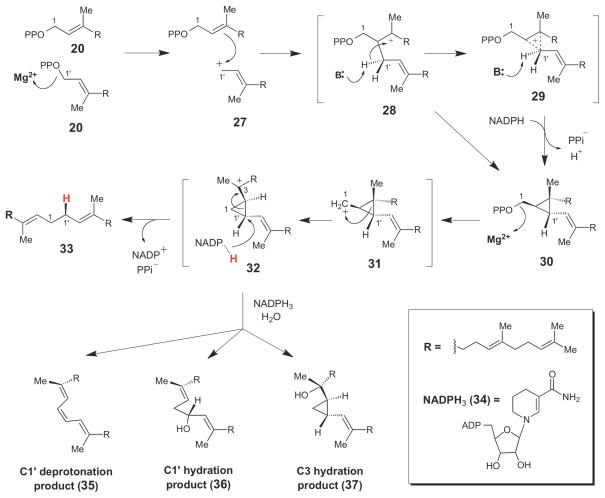
Chemical mechanism of squalene synthase, which proceeds through the stable cyclopropyl intermediate, presqualene pyrophosphate (30).
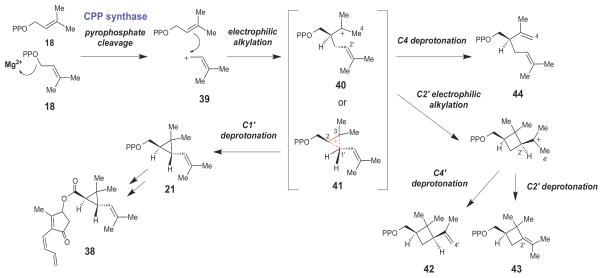
Proposed chemical mechanism for chrysanthemyl pyrophosphate (CPP) synthase, which catalyzes the first step in the biosynthesis of the pyrethrin class of insecticides.
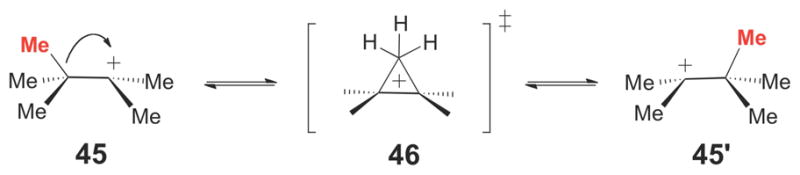
Rearrangement of the tertiary heptyl cation (45) likely involves a protonated cyclopropane transition state (46).
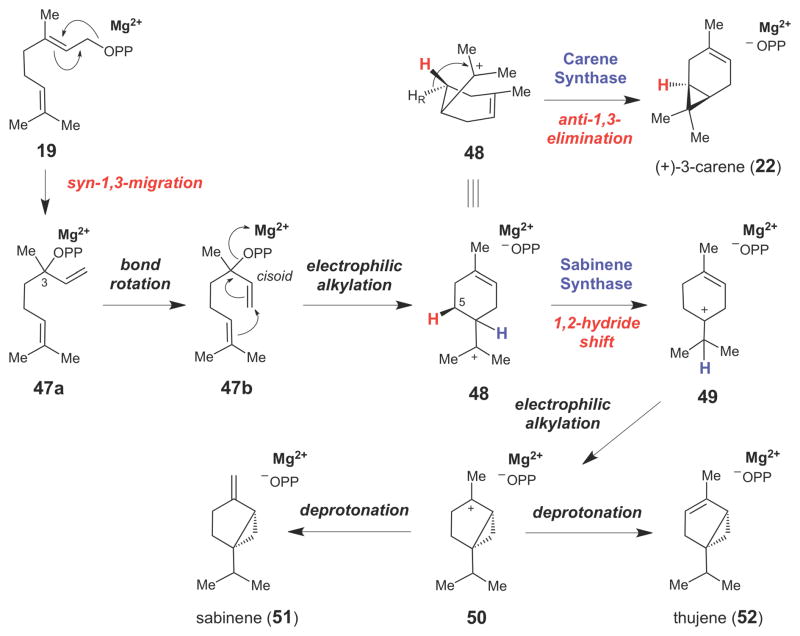
Monoterpene synthase-catalyzed formation of cyclopropane rings in the carene (22), sabinene (51), and thujene (52) families of monoterpenoids.
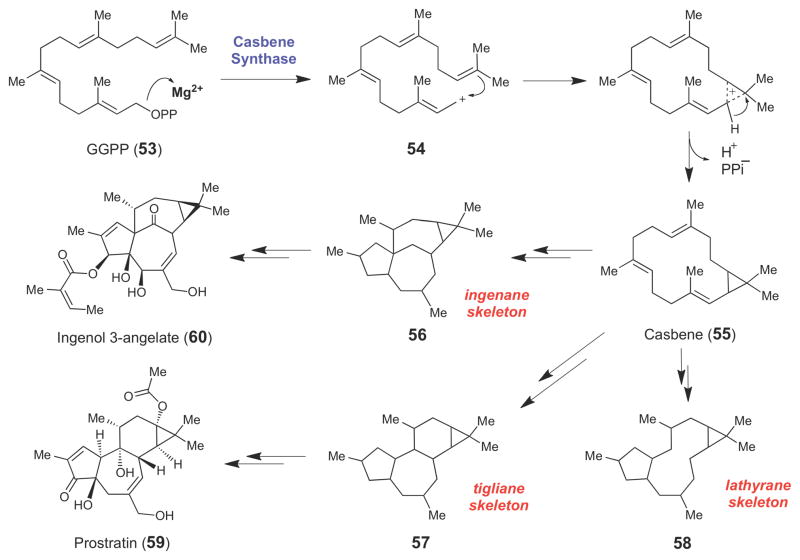
Casbene (55), a diterpene produced from geranylgeranyl pyrophosphate (53) by casbene synthase, is the likely intermediate for the ingenane (56), tigliane (57), and lathyrane (58) families of diterpenoid natural products.
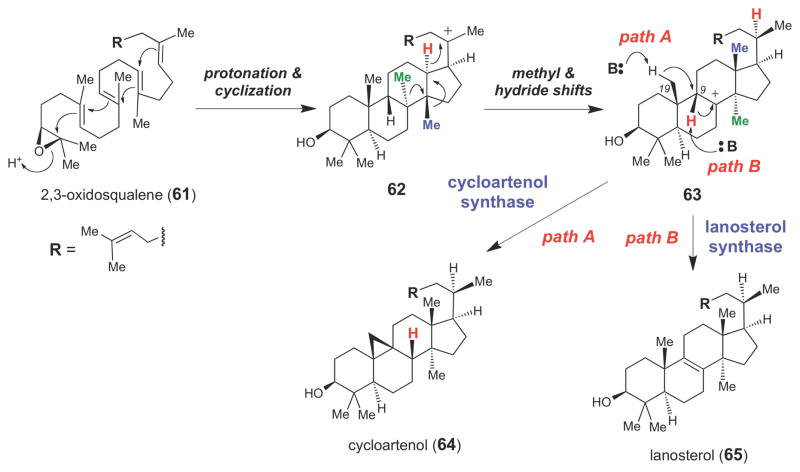
Proposed chemical mechanisms for cycloartenol (64) and lanosterol (65) synthases.
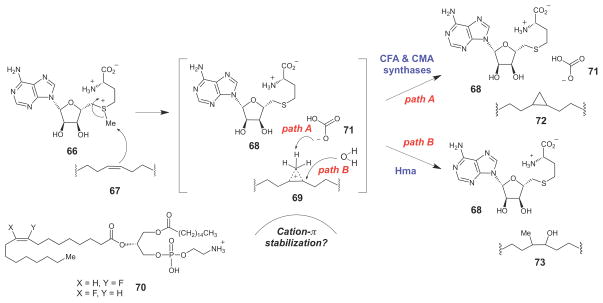
SAM-dependent cyclopropane biosynthesis catalyzed by CFA and CMA synthases.
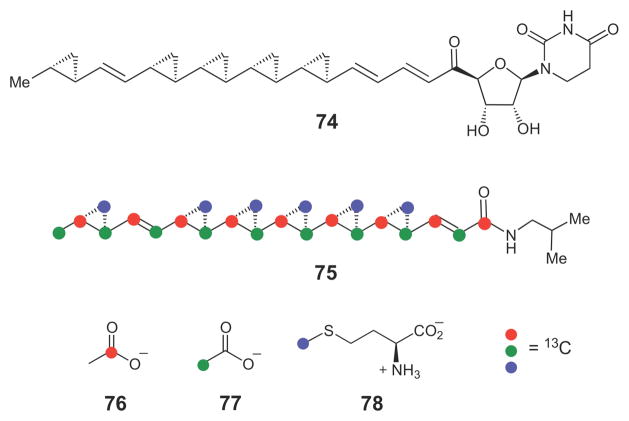
The cyclopropane-containing natural products FR-900848 (74) and U-106305 (75) are likely biosynthesized by a series of insertion of a SAM-derived methylene group into each of the reacting alkene moiety.
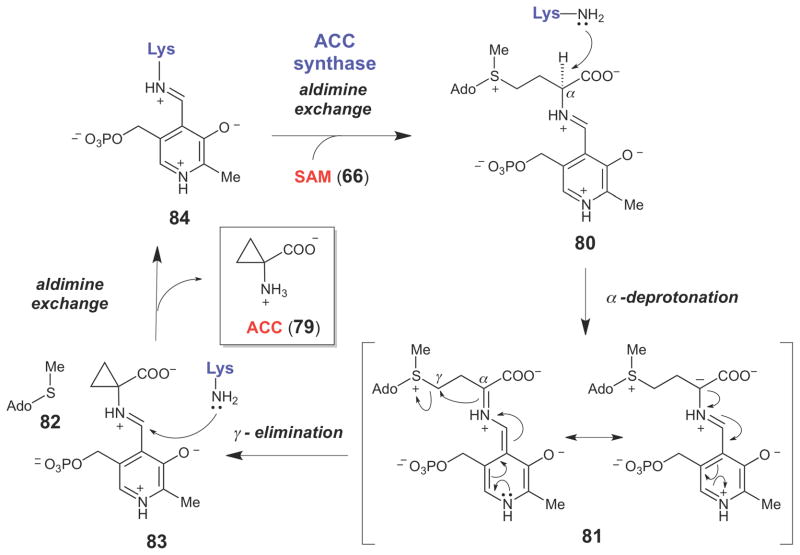
ACC synthase-catalyzed cyclopropane formation, which proceeds from the resonance-stabilized carbanion (81) via an intramolecular SN2 reaction.
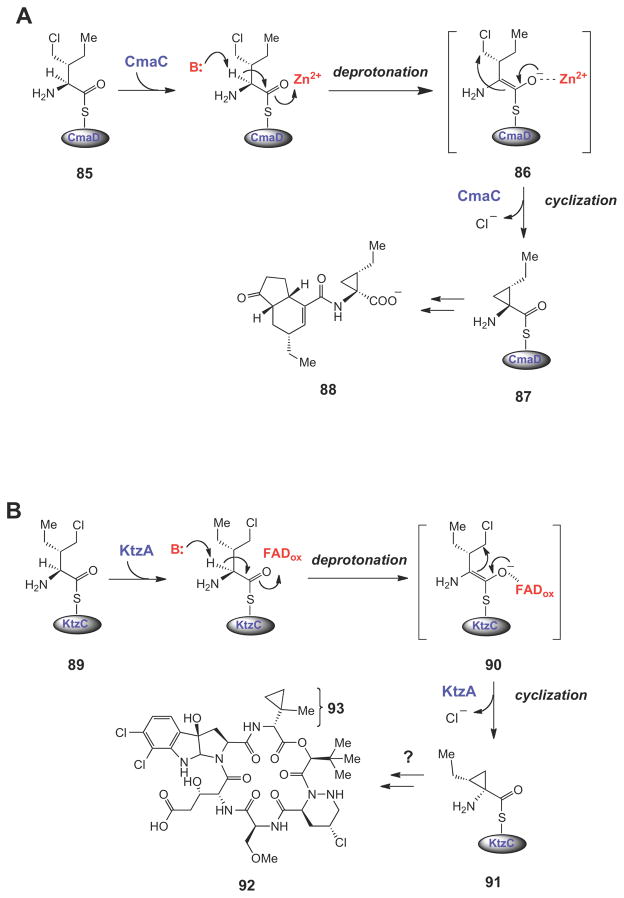
Halogenated carrier protein-linked intermediates serve as the substrates for SN2-like cyclopropane ring formation in the biosynthesis of coronatine (88), kutzneride 2 (92), and curacin A (97). The triggering mechanism for cyclopropane formation in each case is believed to involve a carbanionic intermediate (or transition state).
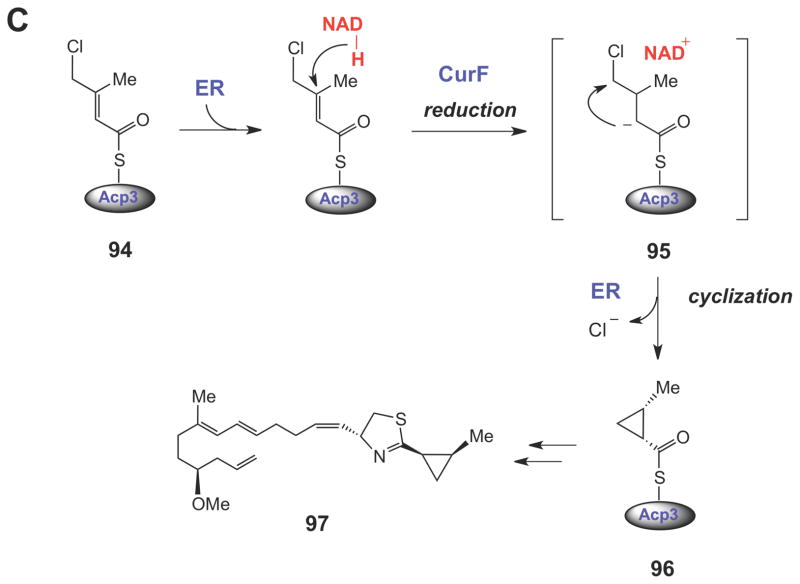
Halogenated carrier protein-linked intermediates serve as the substrates for SN2-like cyclopropane ring formation in the biosynthesis of coronatine (88), kutzneride 2 (92), and curacin A (97). The triggering mechanism for cyclopropane formation in each case is believed to involve a carbanionic intermediate (or transition state).
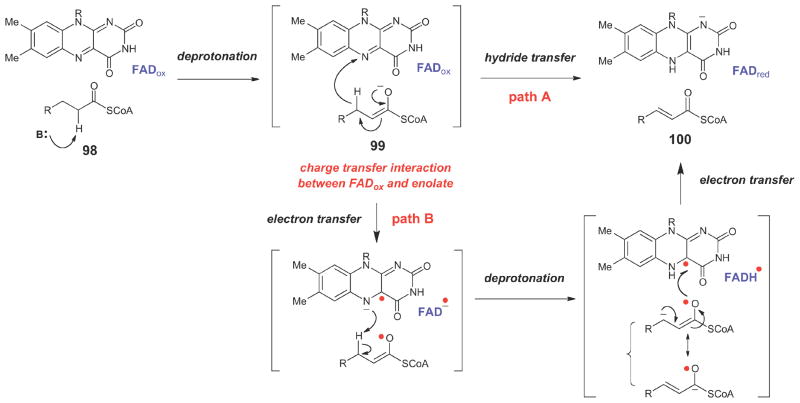
Generalized chemical mechanism for acyl-CoA dehydrogenases.
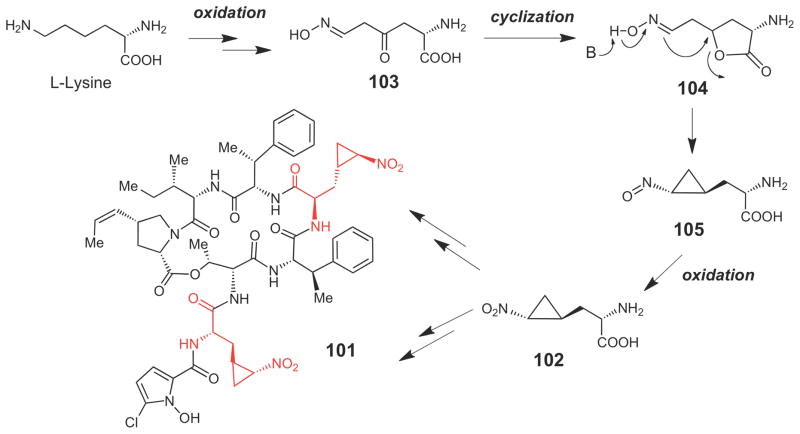
Putative mechanism for cyclopropane formation in hormaomycin (101).
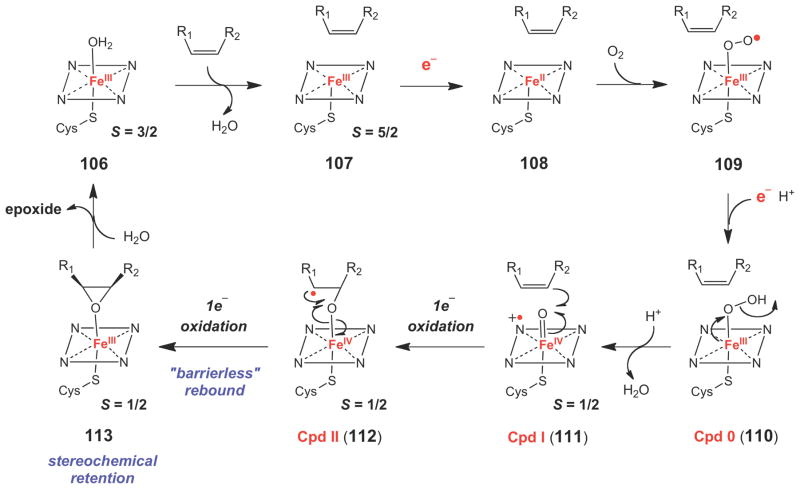
Typical catalytic cycle for a P450-dependent epoxidase.
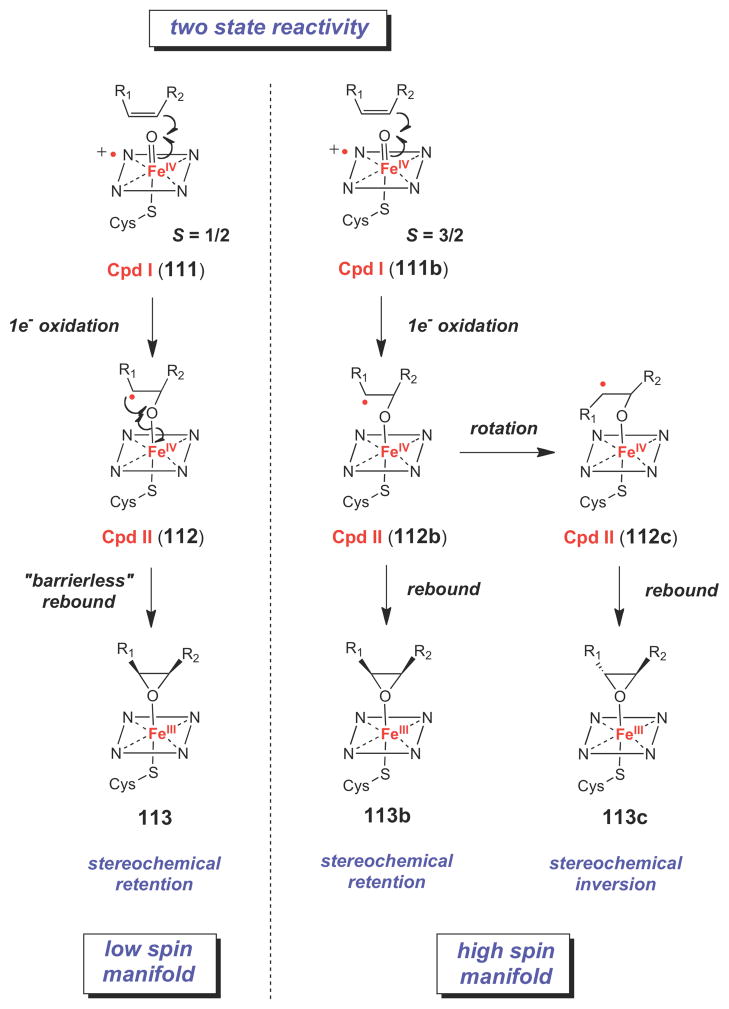
Two-state reactivity model for P450 enzymes.
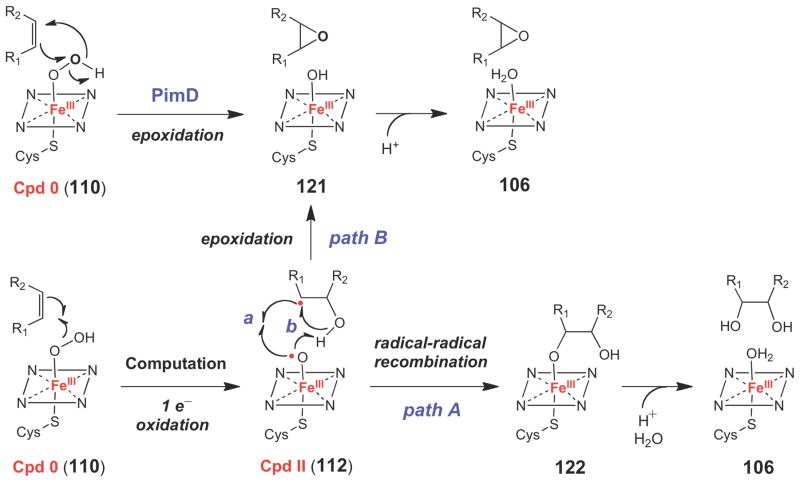
Alternative mechanism of epoxidation by P450 enzymes involving a Cpd 0 species (110).
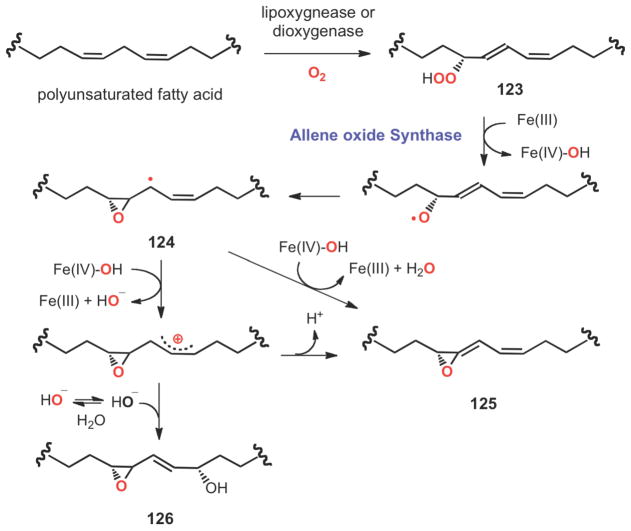
Radical and ionic pathways to allene oxide (125) catalyzed by a catalase-related allene oxide synthase from the cyanobacterium Acaryochloris marina.
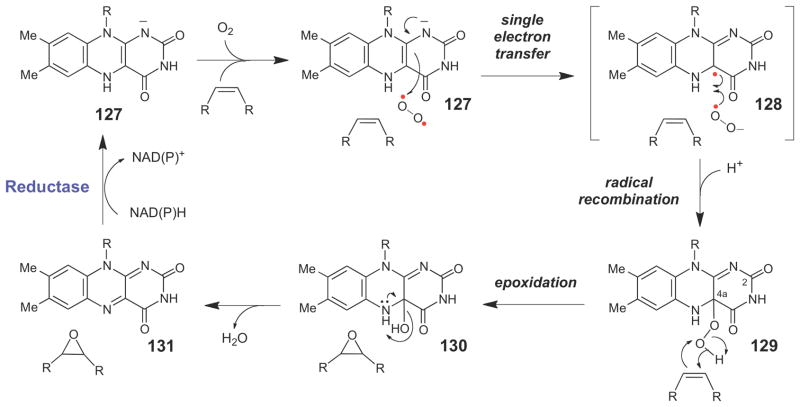
Generalized chemical mechanism for flavin dependent epoxidases.
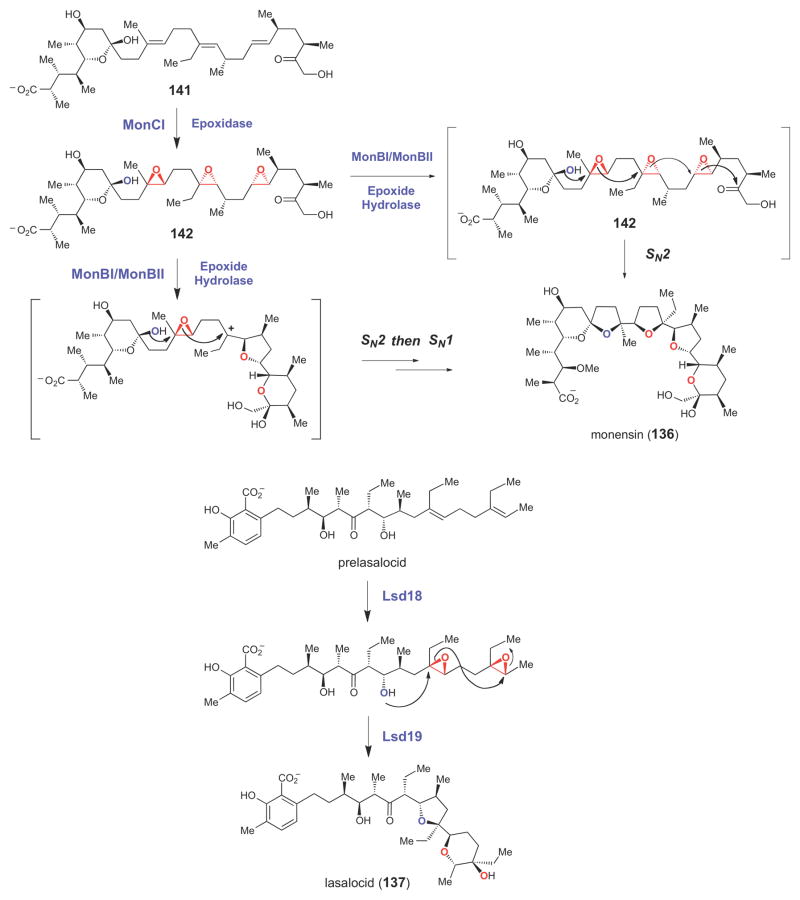
Involvement of epoxide intermediate in the biosynthesis of polyether natural products.
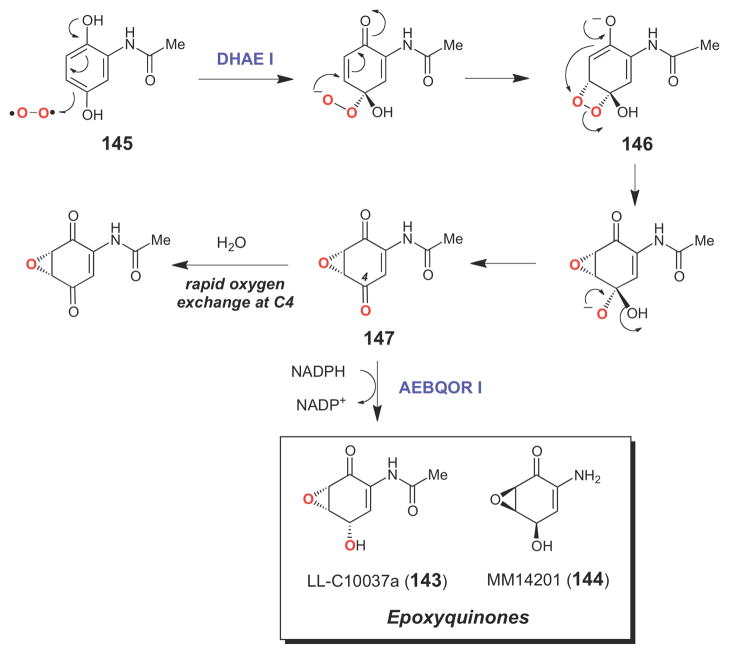
Cofactor-independent epoxidation catalyzed by DHAE I.
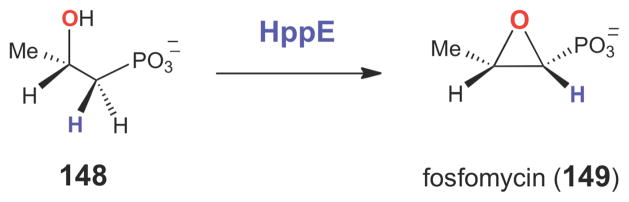
The epoxidation reaction catalyzed by HppE involves the two-electron oxidation of a secondary alcohol.
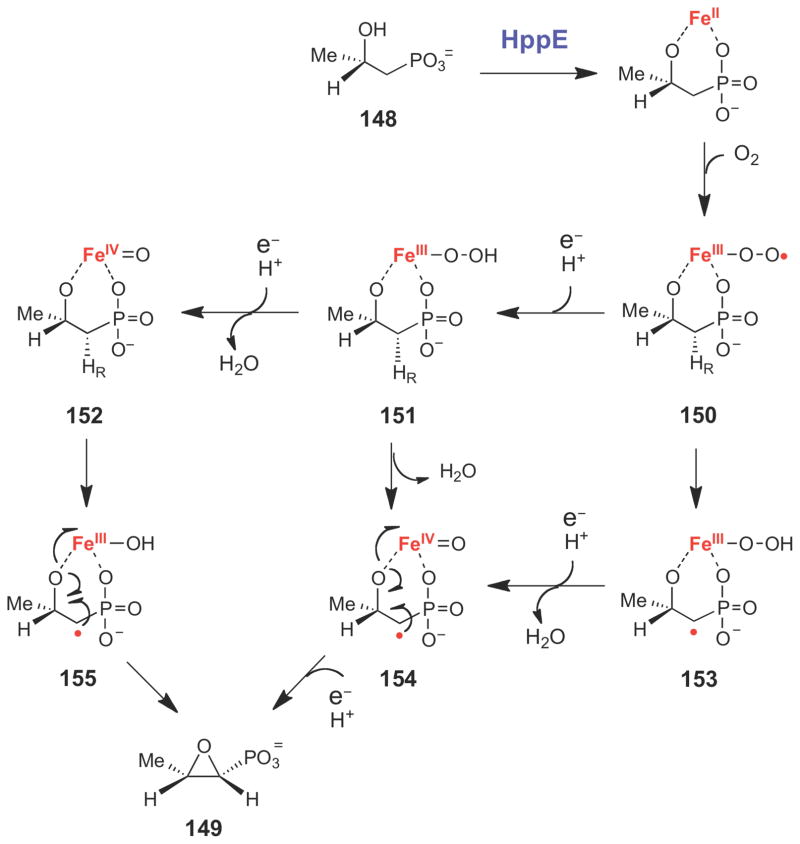
Proposed chemical mechanism for HppE.
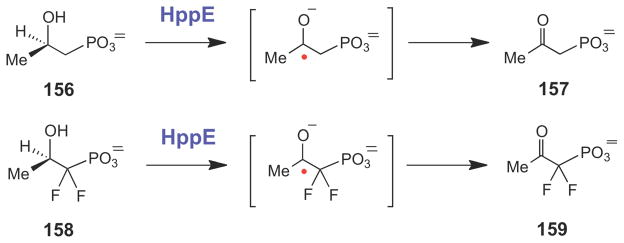
HppE-catalyzed conversion of substrate analogues to the corresponding ketones likely proceeds via hydrogen atom abstraction and ketyl radical intermediates.
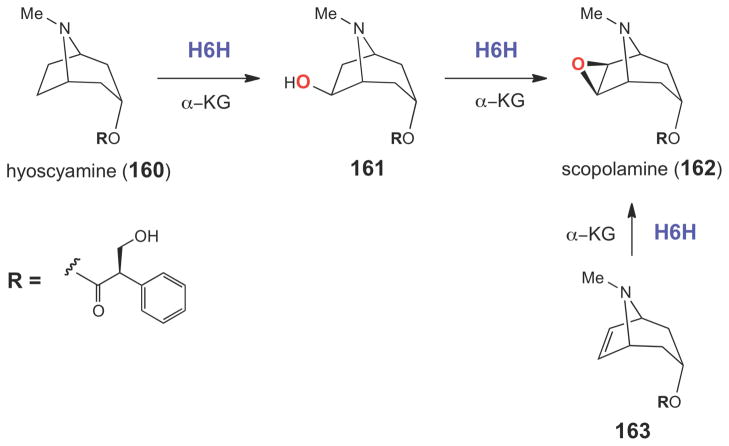
The epoxidation reaction catalyzed by H6H is α-KG-dependent and involves the two-electron oxidation of a secondary alcohol.
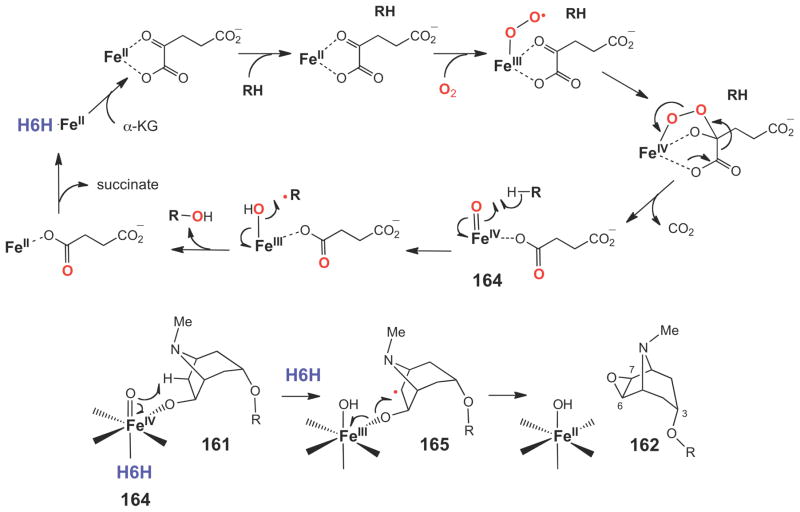
Oxygen activation catalyzed by Fe(II)/α-KG-dependent enzymes and the proposed mechanism of H6H catalyzed two-electron oxidation of a secondary alcohol to generate scopolamine (162).
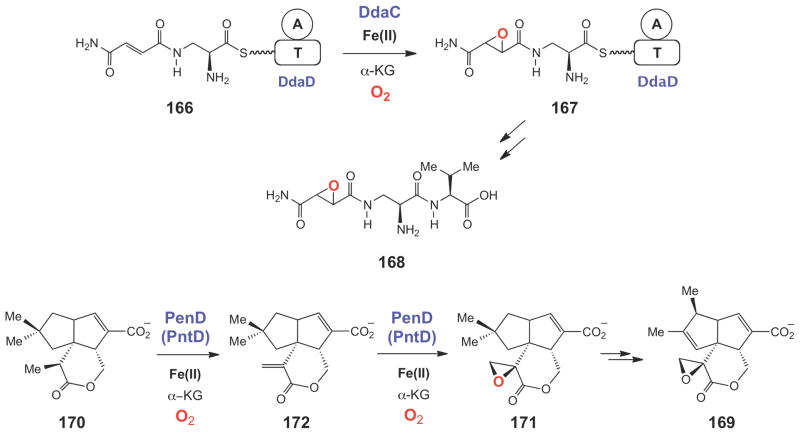
Epoxidation reactions catalyzed by the Fe(II)/α-KG dependent enzymes DdaC and PenD/PntD during Nβ-epoxysuccinamoyl-DAP-Val (168) and pentalenolactone (169) biosynthesis, respectively.
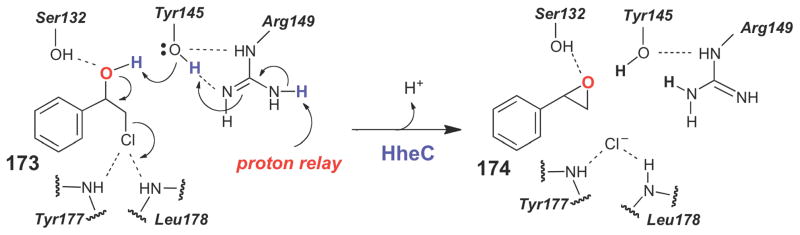
Structure-based mechanism proposed for the haloalcohol dehydrogenase, HheC, which likely catalyzes formation of an epoxide from a 1,2-haloalcohol via an SN2-type reaction.
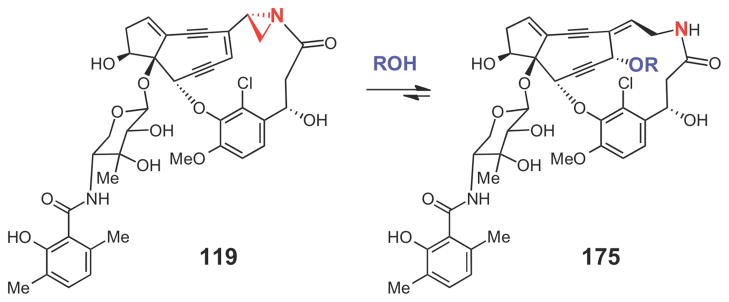
The major compound (175) isolated from the alcohol extract of the tightly bound maduropeptin (119) from MdpA.
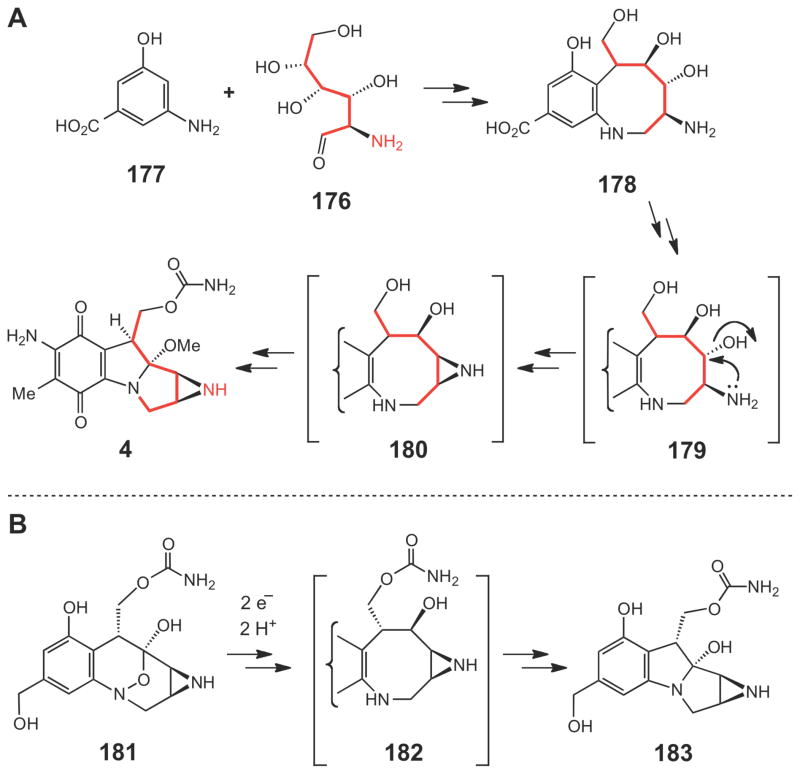
(A) Proposed biosynthetic pathway of mitomycin C starting from glucosamine (176) and AHBA (177). (B) Proposed mechanism of reductive activation of FR66979 (181) via 183 to crosslink DNA.
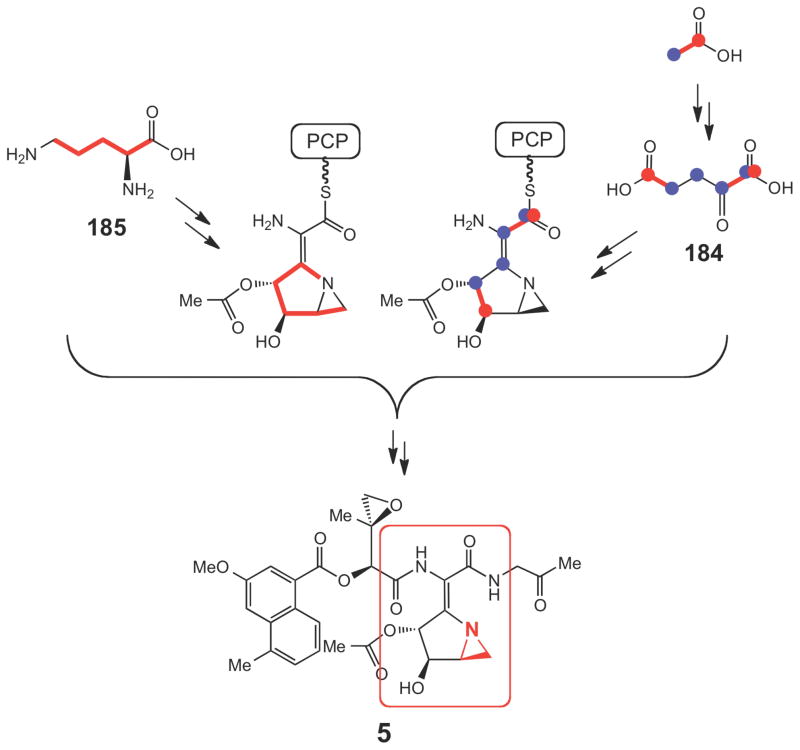
Incorporation of various precursors into the azabicyclic fragment (circled in red) of azinomycin.
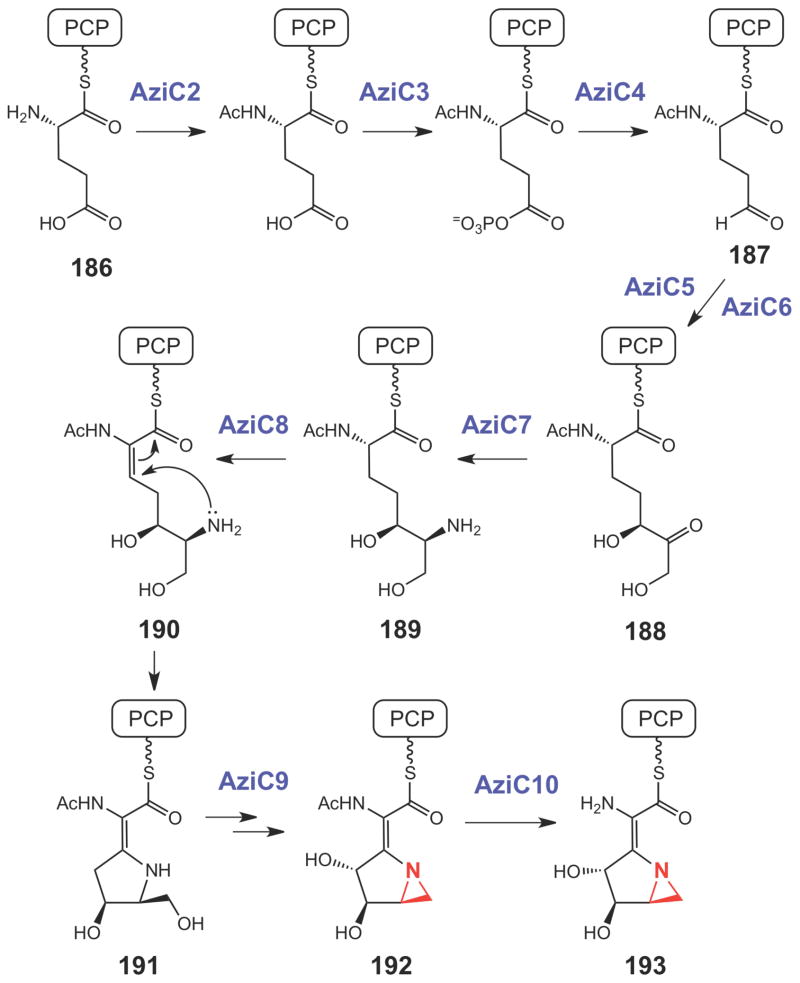
Proposed biosynthetic pathway for the azabicyclic moiety of azinomycin.
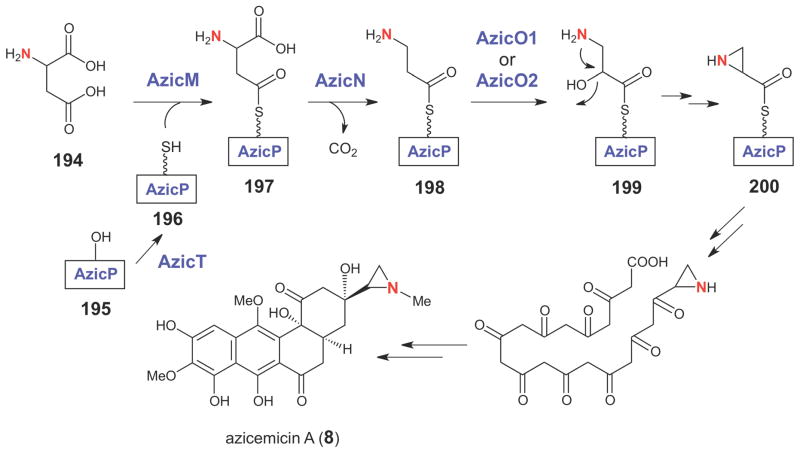
Proposed biosynthetic pathway for aziridine ring formation in the azicemicins.
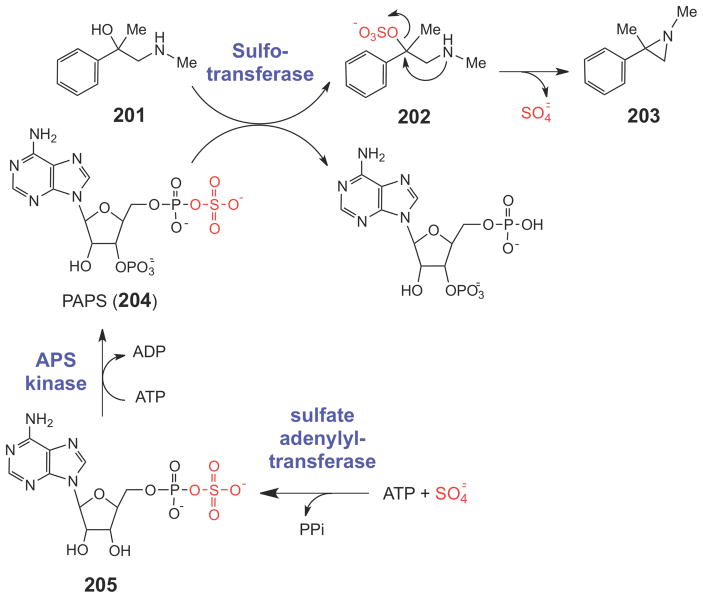
A potential biosynthetic route to the aziridine moiety.
References
-
- Steglich W, Fugmann B, Lang-Fugmann S, editors. Römpp Encyclopedia Natural Products. Georg Thieme Verlag; Stuttgart, Germany: 2000.
-
- Takahashi I, Takahashi K, Ichinura M, Morimoto M, Asano K, Kawamoto I, Tomita F, Nakano H. J Antibiot. 1988;41:1915. - PubMed
-
- Konichi M, Ohkuma H, Matusumoto K, Tsuno T, Kamei H, Miyaki T, Oki T, Kawaguchi H, Van Duyne GD, Clardy J. J Antibiot. 1989;42:1449. - PubMed
-
- Hofle G, Bedorf N, Steinmetz H, Schomburg D, Gerth K, Reichenbach H. Angew Chem Int Ed. 1996;35:1567.
-
- Tomasz M, Palom Y. Pharmacol Therap. 1997;76:73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
