

Primary motor cortex reports efferent control of vibrissa motion on multiple timescales
- PMID: 22017992
- PMCID: PMC3717360
- DOI: 10.1016/j.neuron.2011.09.020
Primary motor cortex reports efferent control of vibrissa motion on multiple timescales
Abstract
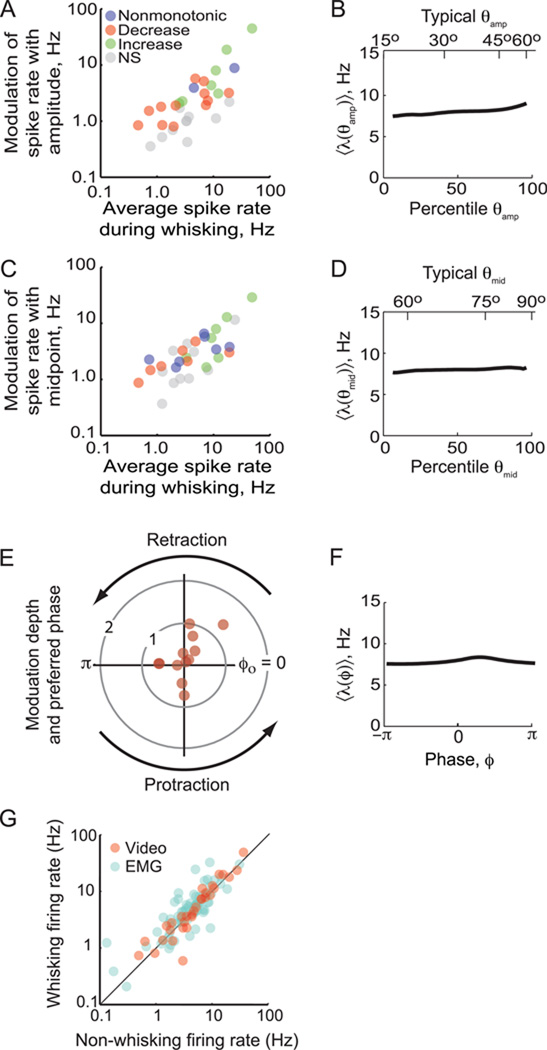
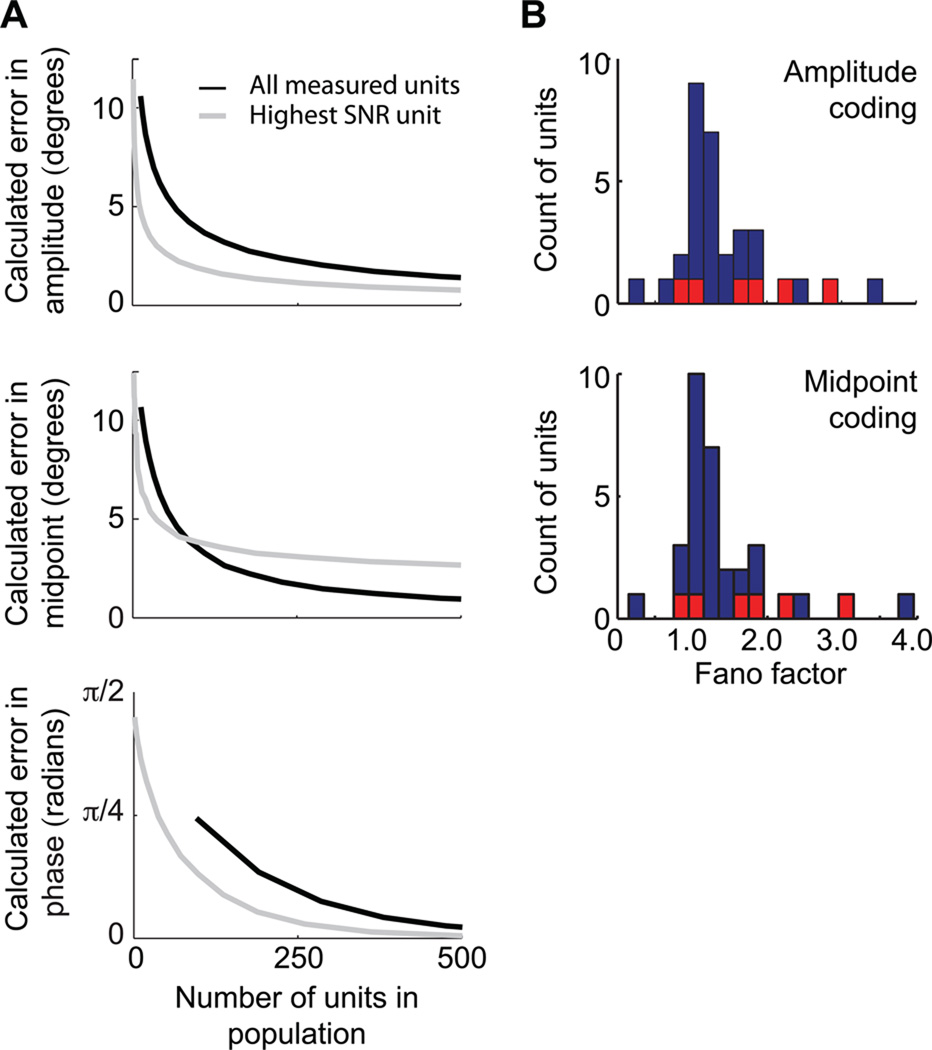
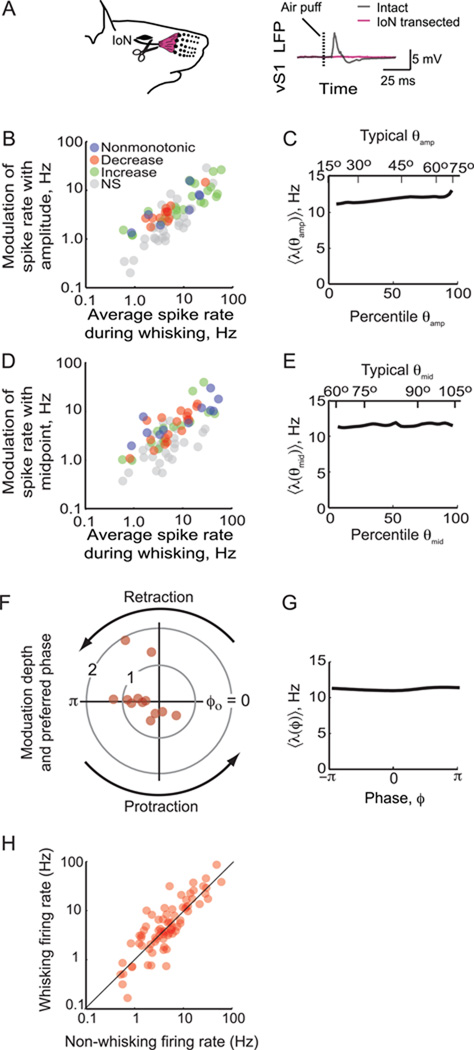
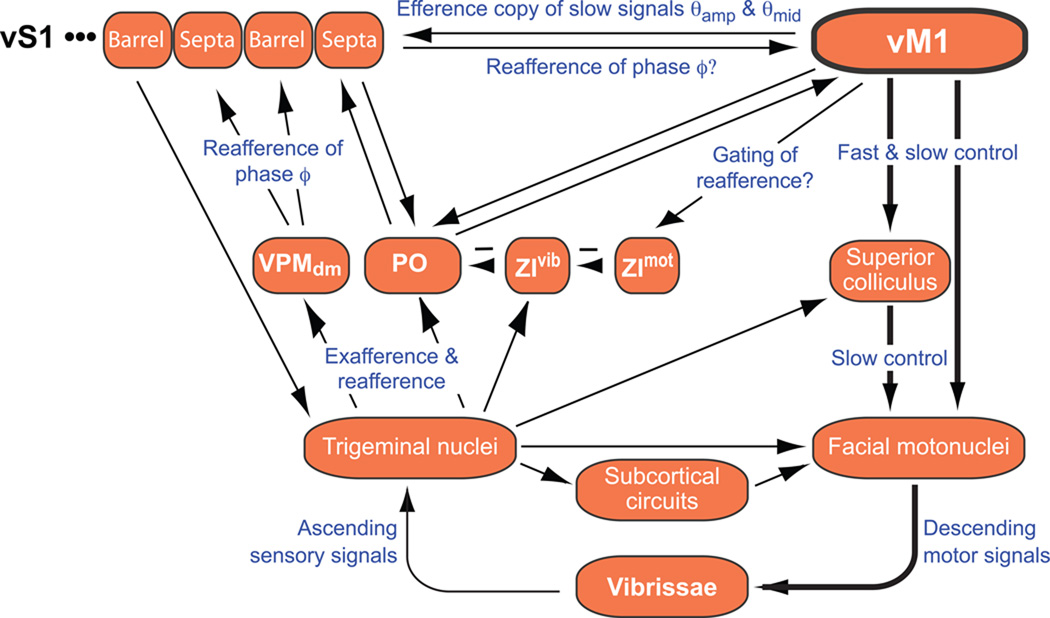
Exploratory whisking in rat is an example of self-generated movement on multiple timescales, from slow variations in the envelope of whisking to the rapid sequence of muscle contractions during a single whisk cycle. We find that, as a population, spike trains of single units in primary vibrissa motor cortex report the absolute angle of vibrissa position. This representation persists after sensory nerve transection, indicating an efferent source. About two-thirds of the units are modulated by slow variations in the envelope of whisking, while relatively few units report rapid changes in position within the whisk cycle. The combined results from this study and past measurements, which show that primary sensory cortex codes the whisking envelope as a motor copy signal, imply that signals present in both sensory and motor cortices are necessary to compute angular coordinates based on vibrissa touch.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
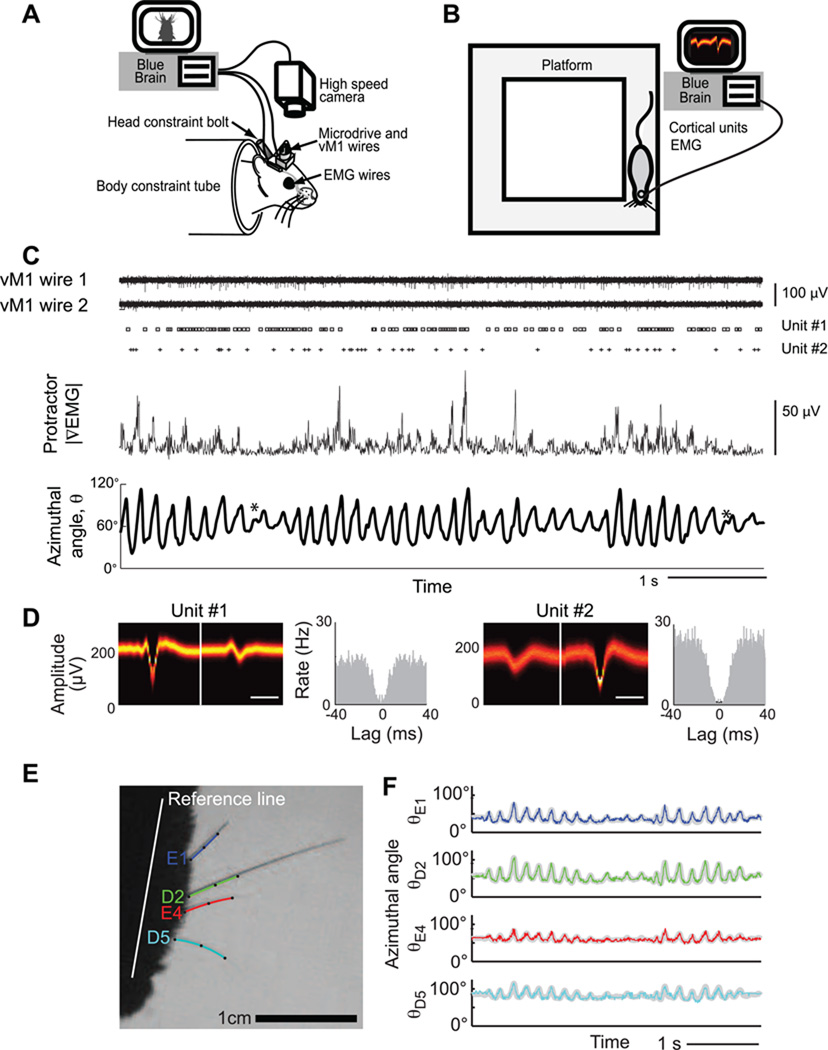
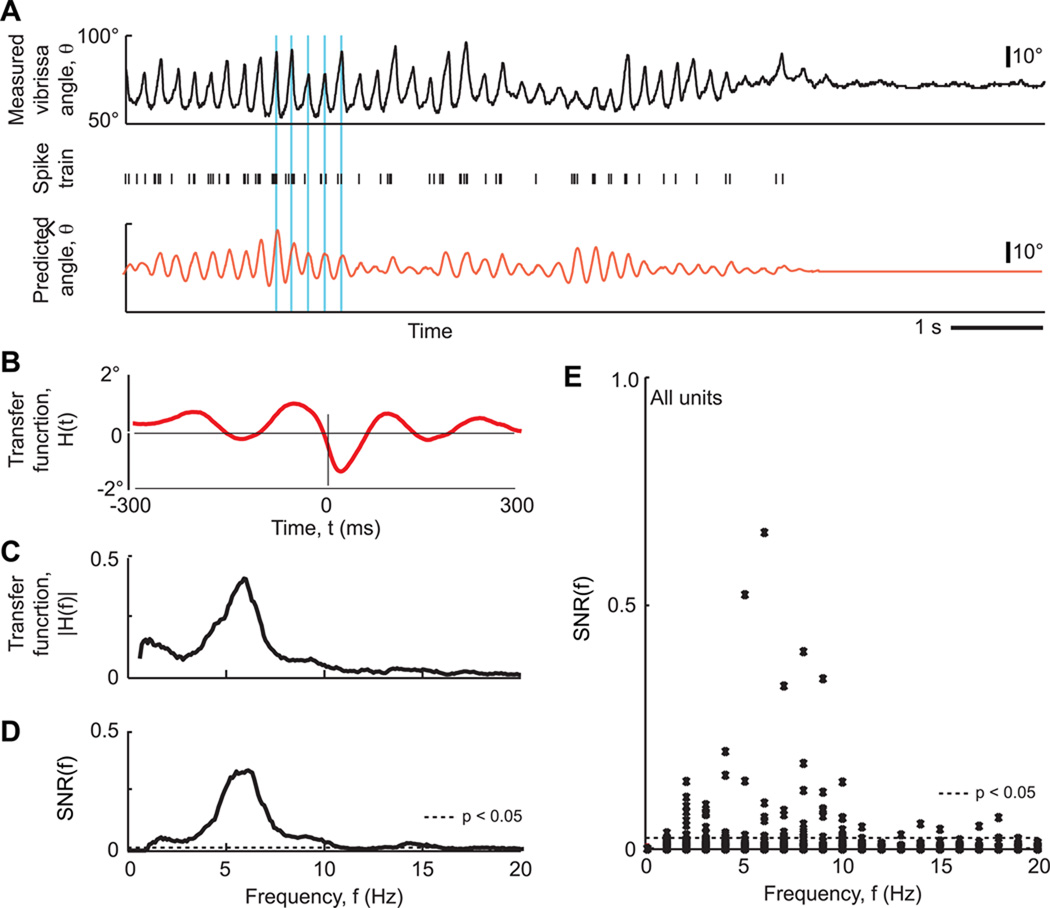
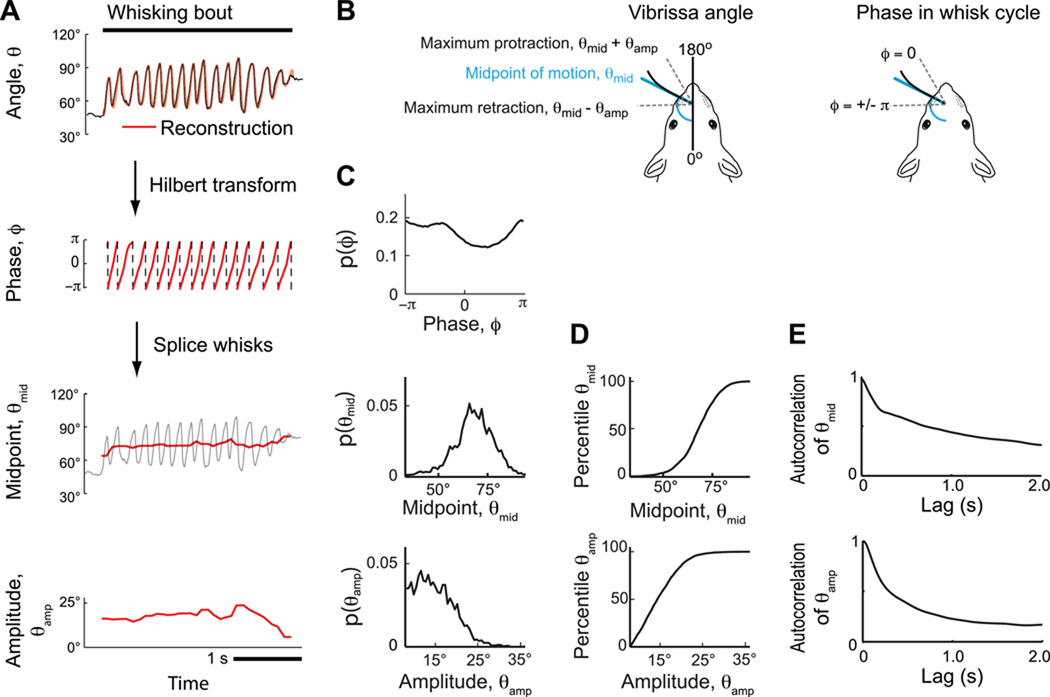
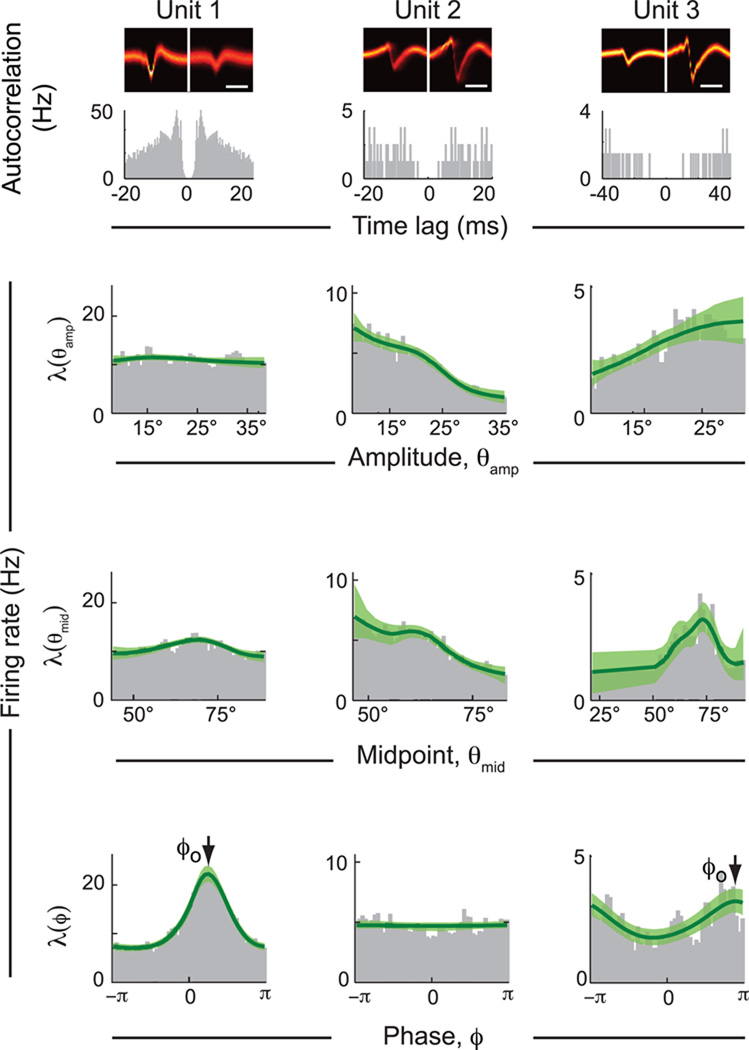
Comment in
-
Movement, confusion, and orienting in frontal cortices.Neuron. 2011 Oct 20;72(2):193-6. doi: 10.1016/j.neuron.2011.10.002. Neuron. 2011. PMID: 22017982
References
-
- Ahissar E, Sosnik R, Haidarliu S. Transformation from temporal to rate coding in a somatosensory thalamocortical pathway. Nature. 2000;406:302–306. - PubMed
-
- Ahrens KF, Kleinfeld D. Current flow in vibrissa motor cortex can phase-lock with exploratory whisking in rat. Journal of Neurophysiology. 2004;92:1700–1707. - PubMed
-
- Arvidsson J, Rice FL. Central projections of primary sensory neurons innervating different parts of the vibrissae follicles and intervibrissal skin on the mystacial pad of the rat. Journal of Comparative Neurology. 1991;309:1–16. - PubMed
-
- Berg RW, Kleinfeld D. Rhythmic whisking by rat: Retraction as well as protraction of the vibrissae is under active muscular control. Journal of Neurophysiology. 2003a;89:104–117. - PubMed
-
- Berg RW, Kleinfeld D. Vibrissa movement elicited by rhythmic electrical microstimuation to motor cortex in the aroused rat mimics exploratory whisking. Journal of Neurophysiology. 2003b;90:2950–2963. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
