

Microbiome and malignancy
- PMID: 22018233
- PMCID: PMC3264051
- DOI: 10.1016/j.chom.2011.10.003
Microbiome and malignancy
Abstract
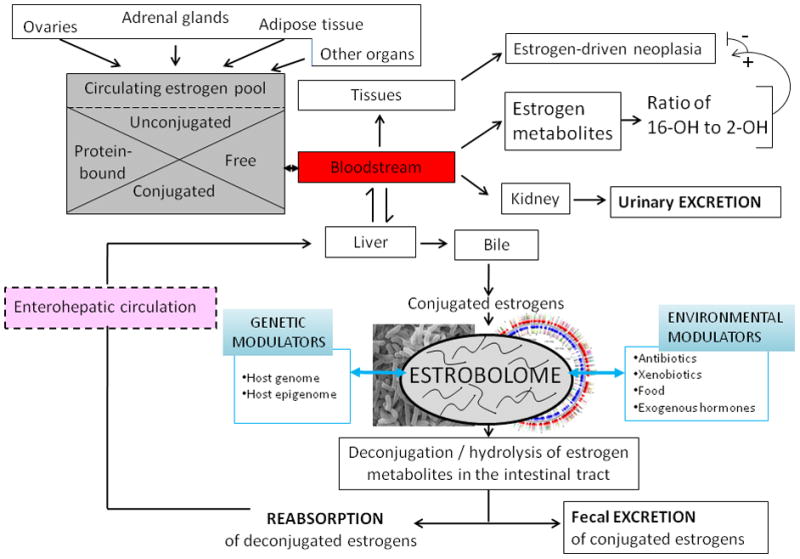
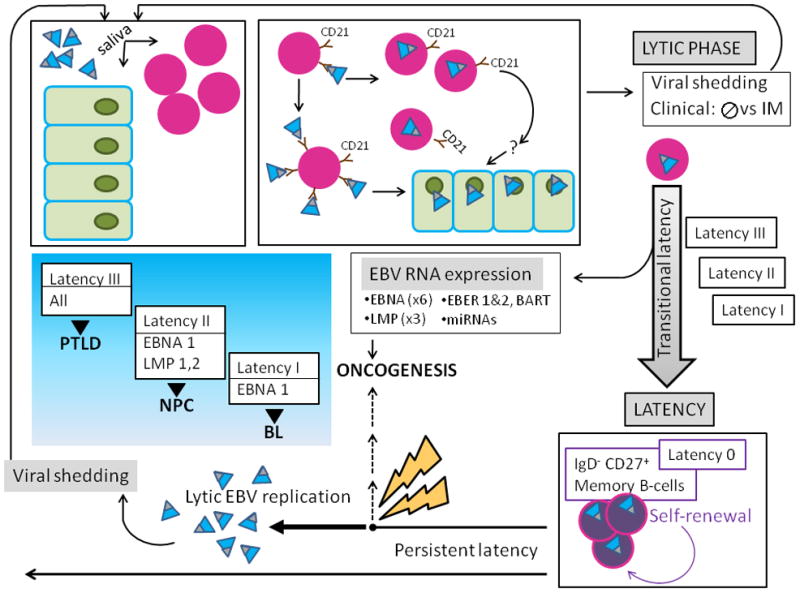
Current knowledge is insufficient to explain why only a proportion of individuals exposed to environmental carcinogens or carrying a genetic predisposition to cancer develop disease. Clearly, other factors must be important, and one such element that has recently received attention is the human microbiome, the residential microbes including Bacteria, Archaea, Eukaryotes, and viruses that colonize humans. Here, we review principles and paradigms of microbiome-related malignancy, as illustrated by three specific microbial-host interactions. We review the effects of the microbiota on local and adjacent neoplasia, present the estrobolome model of distant effects, and discuss the complex interactions with a latent virus leading to malignancy. These are separate facets of a complex biology interfacing all the microbial species we harbor from birth onward toward early reproductive success and eventual senescence.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
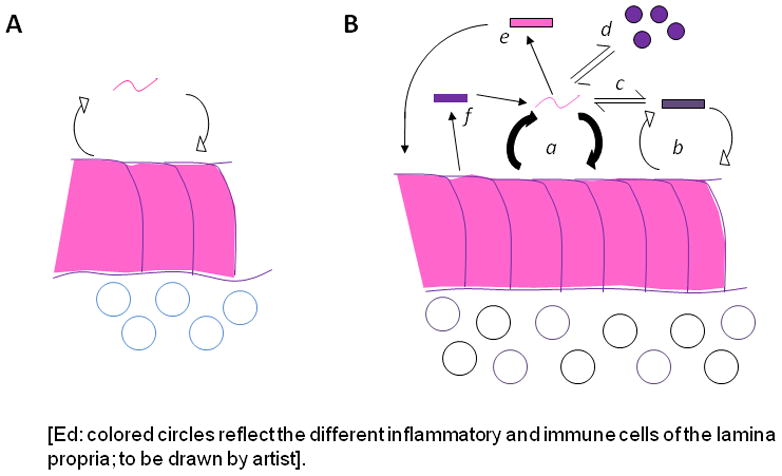
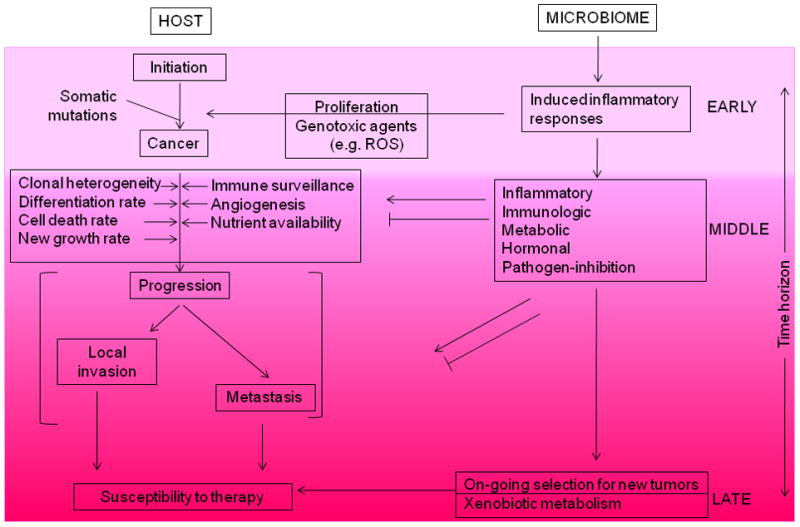
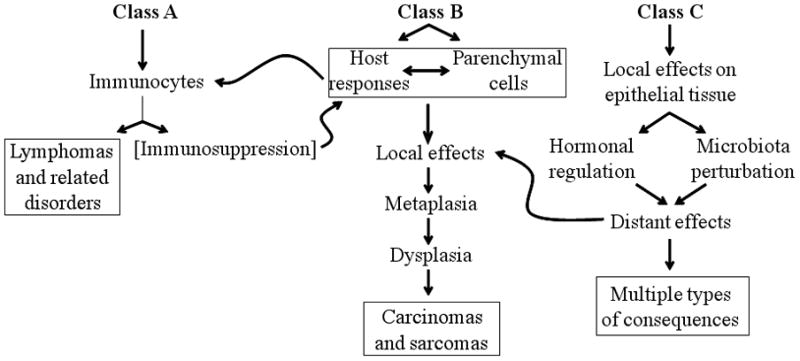
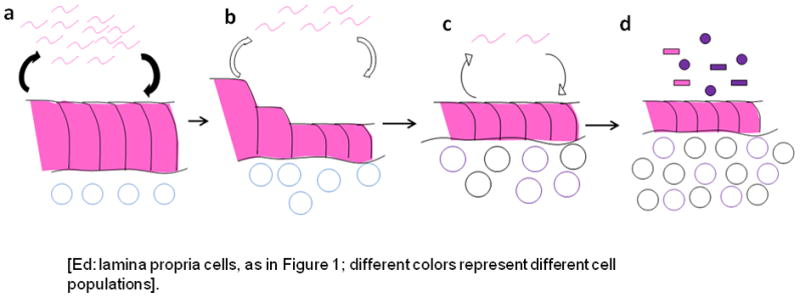
References
-
- Adlercreutz H. Studies on oestrogen excretion in human bile. Acta Endocrinol Suppl (Copenh) 1962;42(Suppl 72):1–220. - PubMed
-
- Adlercreutz H, Jarvenpaa P. Assay of estrogens in human feces. J Steroid Biochem. 1982;17:639–645. - PubMed
-
- Adlercreutz H, Martin F. Biliary excretion and intestinal metabolism of progesterone and estrogens in man. J Steroid Biochem. 1980;13:231–244. - PubMed
-
- Adlercreutz H, Martin F, Jarvenpaa P, Fotsis T. Steroid absorption and enterohepatic recycling. Contraception. 1979;20:201–223. - PubMed
-
- Adlercreutz H, Martin F, Lindstrom B. Gas chromatographic and mass spectrometric studies on oestrogens in bile--2. Men and non-pregnant women. J Steroid Biochem. 1978;9:1197–1205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
