

Differential expression of Ly6C and T-bet distinguish effector and memory Th1 CD4(+) cell properties during viral infection
- PMID: 22018471
- PMCID: PMC3444169
- DOI: 10.1016/j.immuni.2011.08.016
Differential expression of Ly6C and T-bet distinguish effector and memory Th1 CD4(+) cell properties during viral infection
Abstract
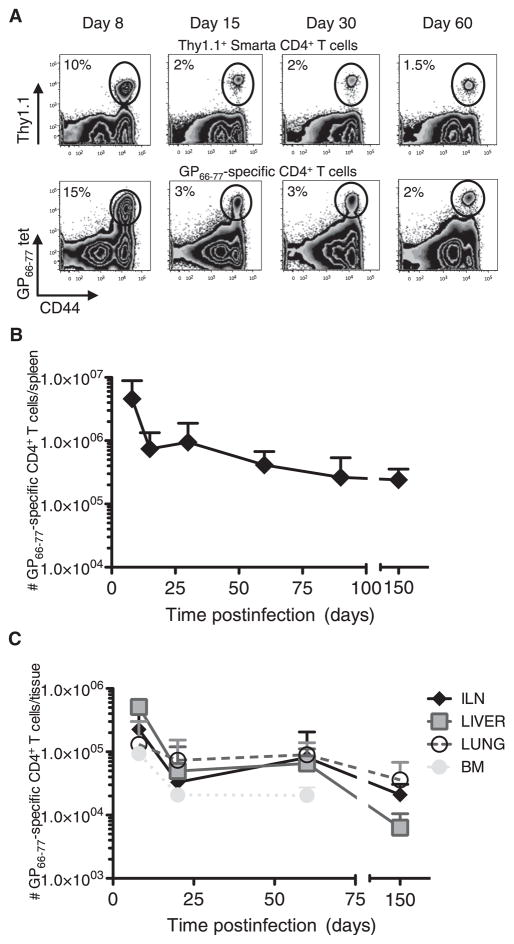
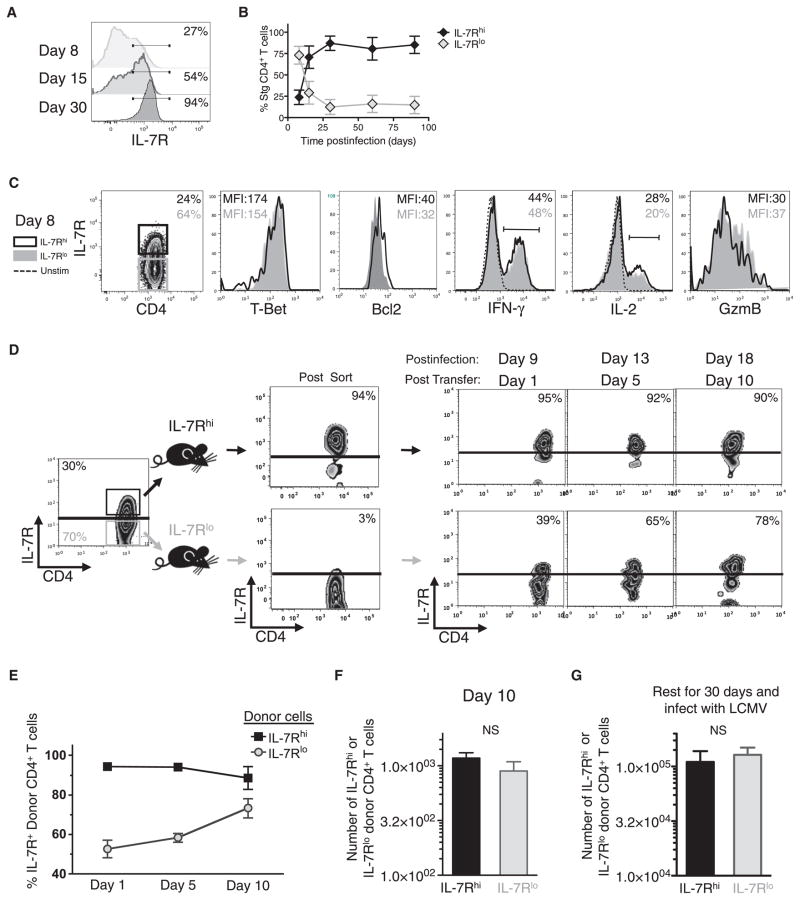
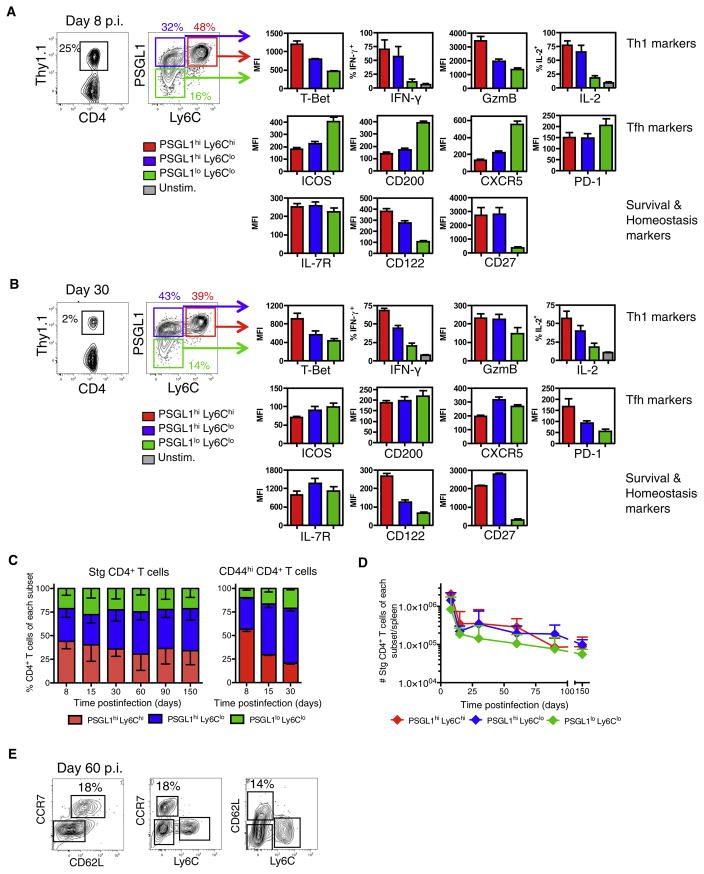
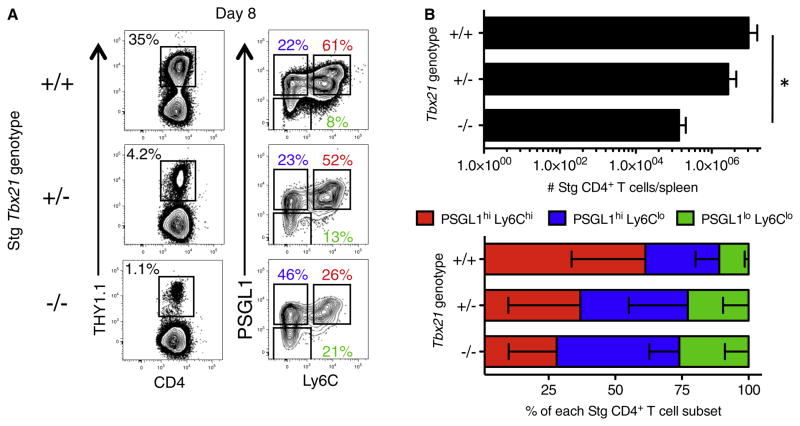
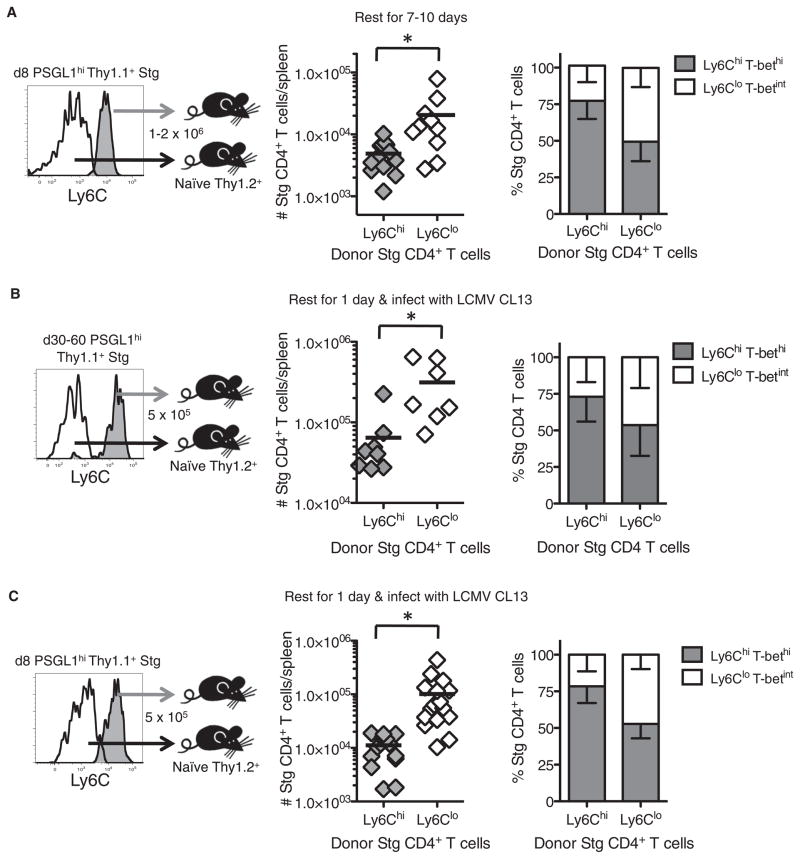
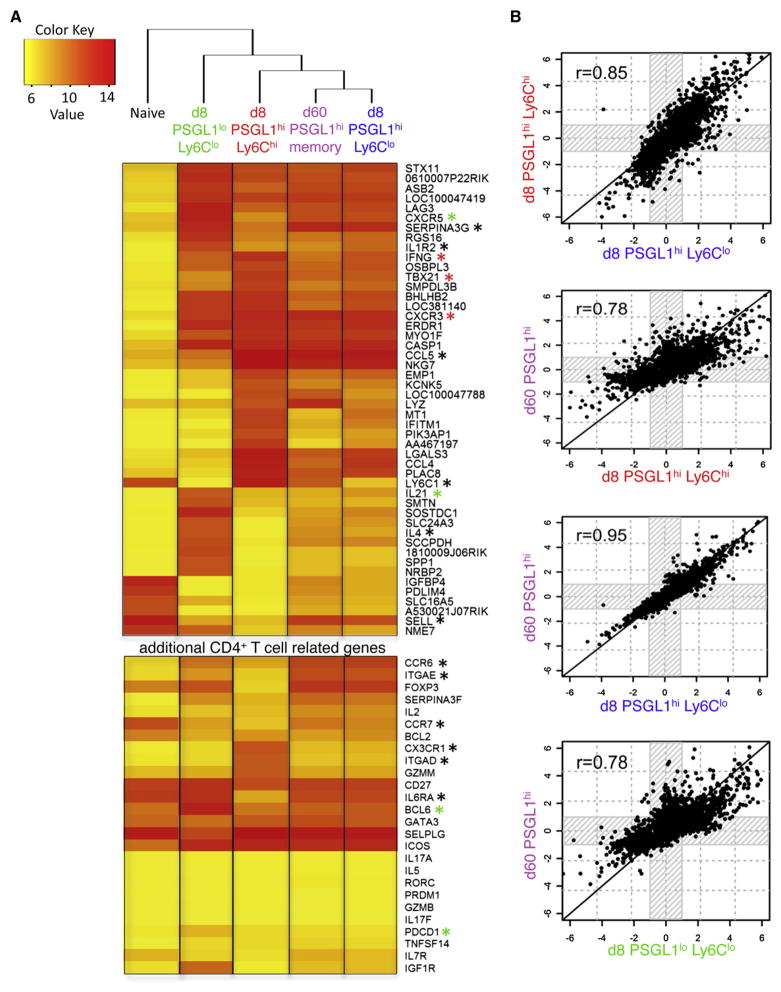
CD4(+) T cells differentiate into multiple effector types, but it is unclear how they form memory T cells during infection in vivo. Profiling virus-specific CD4(+) T cells revealed that effector cells with T helper 1 (Th1) or T follicular helper (Tfh) cell characteristics differentiated into memory cells, although expression of Tfh cell markers declined over time. In contrast to virus-specific effector CD8(+) T cells, increased IL-7R expression was not a reliable marker of CD4(+) memory precursor cells. However, decreased Ly6C and T-bet (Tbx21) expression distinguished a subset of Th1 cells that displayed greater longevity and proliferative responses to secondary infection. Moreover, the gene expression profile of Ly6C(lo)T-bet(int) Th1 effector cells was virtually identical to mature memory CD4(+) T cells, indicating early maturation of memory CD4(+) T cell features in this subset during acute viral infection. This study provides a framework for memory CD4(+) T cell development after acute viral infection.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Memory: the incomplete unhappening of differentiation.Immunity. 2011 Oct 28;35(4):496-8. doi: 10.1016/j.immuni.2011.10.001. Immunity. 2011. PMID: 22035843
References
-
- Bamezai A. Mouse Ly-6 proteins and their extended family: markers of cell differentiation and regulators of cell signaling. Arch Immunol Ther Exp (Warsz) 2004;52:255–266. - PubMed
-
- Blander JM, Sant’Angelo DB, Bottomly K, Janeway CA., Jr Alteration at a single amino acid residue in the T cell receptor alpha chain complementarity determining region 2 changes the differentiation of naive CD4 T cells in response to antigen from T helper cell type 1 (Th1) to Th2. J Exp Med. 2000;191:2065–2074. - PMC - PubMed
-
- De Riva A, Bourgeois C, Kassiotis G, Stockinger B. Noncognate interaction with MHC class II molecules is essential for maintenance of T cell metabolism to establish optimal memory CD4 T cell function. J Immunol. 2007;178:5488–5495. - PubMed
-
- Fazilleau N, Eisenbraun MD, Malherbe L, Ebright JN, Pogue-Caley RR, McHeyzer-Williams LJ, McHeyzer-Williams MG. Lymphoid reservoirs of antigen-specific memory T helper cells. Nat Immunol. 2007;8:753–761. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
