

The first bromodomain of the testis-specific double bromodomain protein Brdt is required for chromocenter organization that is modulated by genetic background
- PMID: 22020252
- PMCID: PMC3217133
- DOI: 10.1016/j.ydbio.2011.10.005
The first bromodomain of the testis-specific double bromodomain protein Brdt is required for chromocenter organization that is modulated by genetic background
Abstract
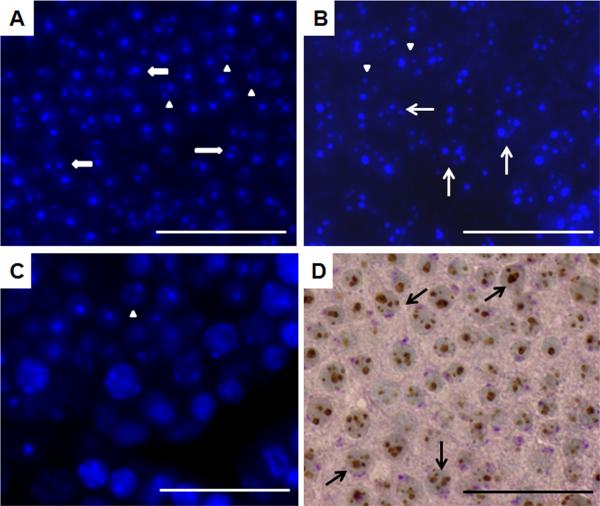
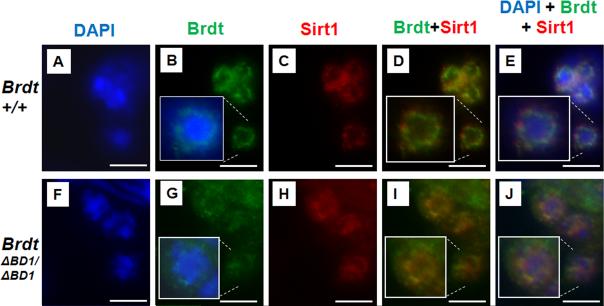
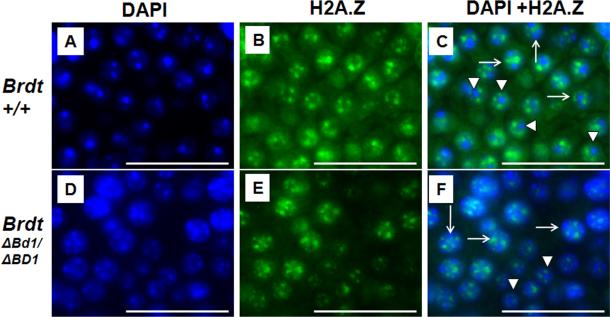
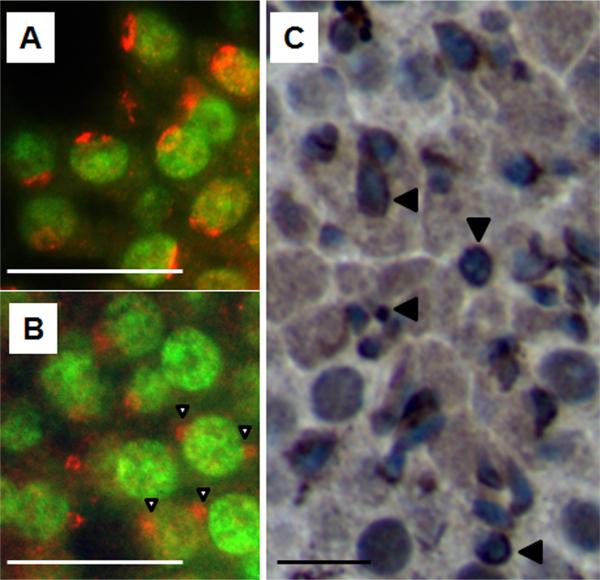
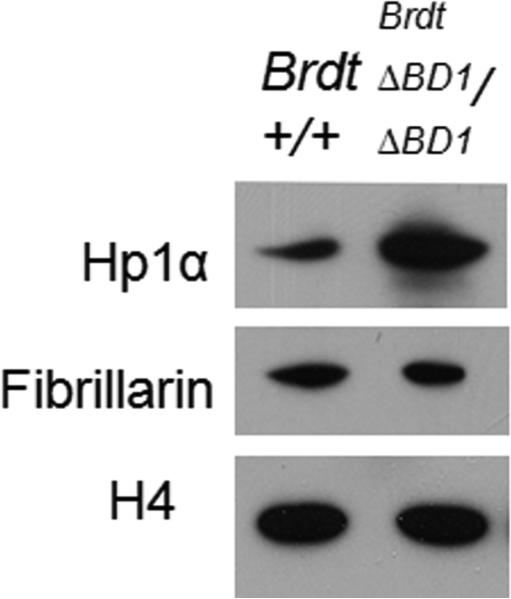
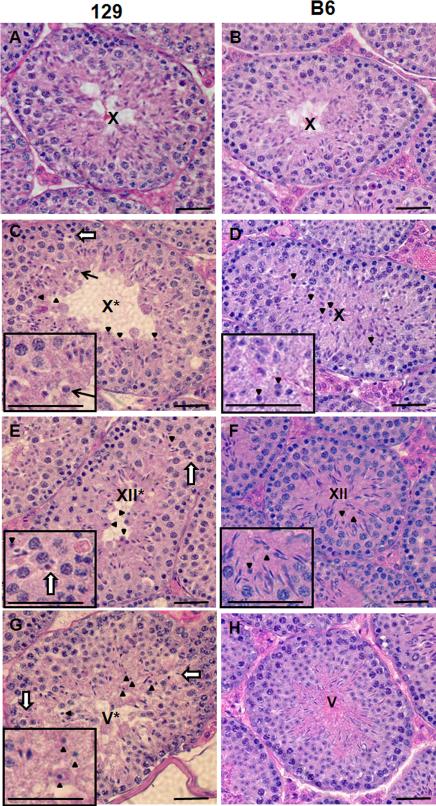
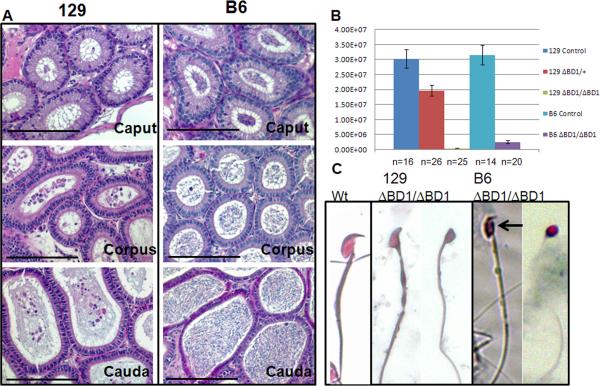
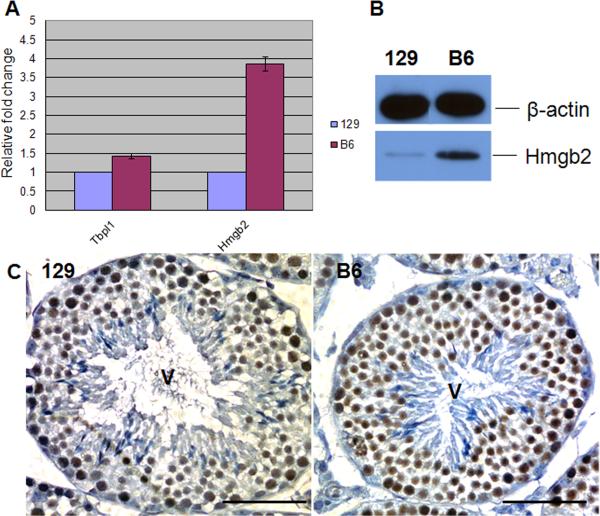
Mice homozygous for a mutation (Brdt(∆BD1/∆BD1)) lacking the first bromodomain of Brdt, a testis-specific member of the BET family of double-bromodomain containing proteins, are sterile and exhibit profound defects in chromatin remodeling during spermiogenesis. We have now observed that a prominent feature of the aberrant spermatid nuclei is a fragmented chromocenter, a structure comprised of peri-centromeric heterochromatin. There was a concomitant increase in the levels of heterochromatin protein 1 alpha (Hp1α), suggesting that the presence of multiple chromocenters was correlated with a spread of heterochromatin beyond the normal centromeric region. Brdt protein was normally present throughout the nucleus but was excluded from the chromocenter. A more densely staining region of Brdt protein appeared to separate sirtuin 1 (Sirt1) protein from contact with the chromocenter. Although still nuclear, this unique localization of Brdt protein was lost in Brdt(∆BD1/∆BD1) mutant spermatids and Brdt and Sirt1 overlapped around the chromocenters. There was also ectopic localization of the H1 histone family, member N, testis-specific (H1fnt) protein in Brdt(∆BD1/∆BD1) round spermatids, which may be linked to the previously reported loss of polarized localization of peri-nuclear heterochromatin foci. The extent of chromocenter fragmentation was more severe and penetrant in mutant testes on a pure 129Sv/Ev as compared to a pure C57Bl/6 background. Indeed, all aspects of the mutant phenotype were more severe on the 129Sv/Ev background. Contrary to previous studies in genetic models where fragmented chromocenters were observed in spermatids, the Brdt(∆BD1/∆BD1) mutant spermatids do not undergo apoptosis (on either background). These observations suggest that the first bromodomain of Brdt is critical in the formation and/or maintenance of an intact chromocenter and implicate this structure in proper remodeling of the chromatin architecture of the sperm head.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–4. - PubMed
-
- Catena R, Ronfani L, Sassone-Corsi P, Davidson I. Changes in intranuclear chromatin architecture induce bipolar nuclear localization of histone variant H1T2 in male haploid spermatids. Dev Biol. 2006;296:231–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
