

miR-34 miRNAs provide a barrier for somatic cell reprogramming
- PMID: 22020437
- PMCID: PMC3541684
- DOI: 10.1038/ncb2366
miR-34 miRNAs provide a barrier for somatic cell reprogramming
Abstract
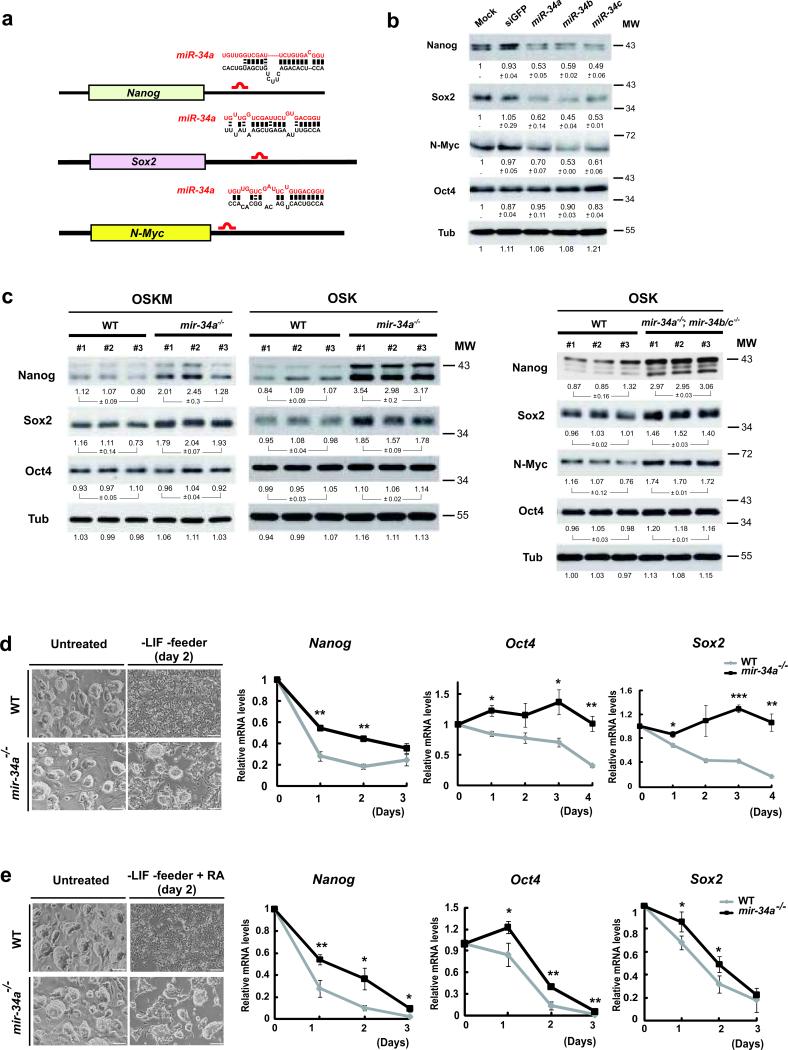
Somatic reprogramming induced by defined transcription factors is a low-efficiency process that is enhanced by p53 deficiency. So far, p21 is the only p53 target shown to contribute to p53 repression of iPSC (induced pluripotent stem cell) generation, indicating that additional p53 targets may regulate this process. Here, we demonstrate that miR-34 microRNAs (miRNAs), particularly miR-34a, exhibit p53-dependent induction during reprogramming. Mir34a deficiency in mice significantly increased reprogramming efficiency and kinetics, with miR-34a and p21 cooperatively regulating somatic reprogramming downstream of p53. Unlike p53 deficiency, which enhances reprogramming at the expense of iPSC pluripotency, genetic ablation of Mir34a promoted iPSC generation without compromising self-renewal or differentiation. Suppression of reprogramming by miR-34a was due, at least in part, to repression of pluripotency genes, including Nanog, Sox2 and Mycn (also known as N-Myc). This post-transcriptional gene repression by miR-34a also regulated iPSC differentiation kinetics. miR-34b and c similarly repressed reprogramming; and all three miR-34 miRNAs acted cooperatively in this process. Taken together, our findings identified miR-34 miRNAs as p53 targets that play an essential role in restraining somatic reprogramming.
Figures
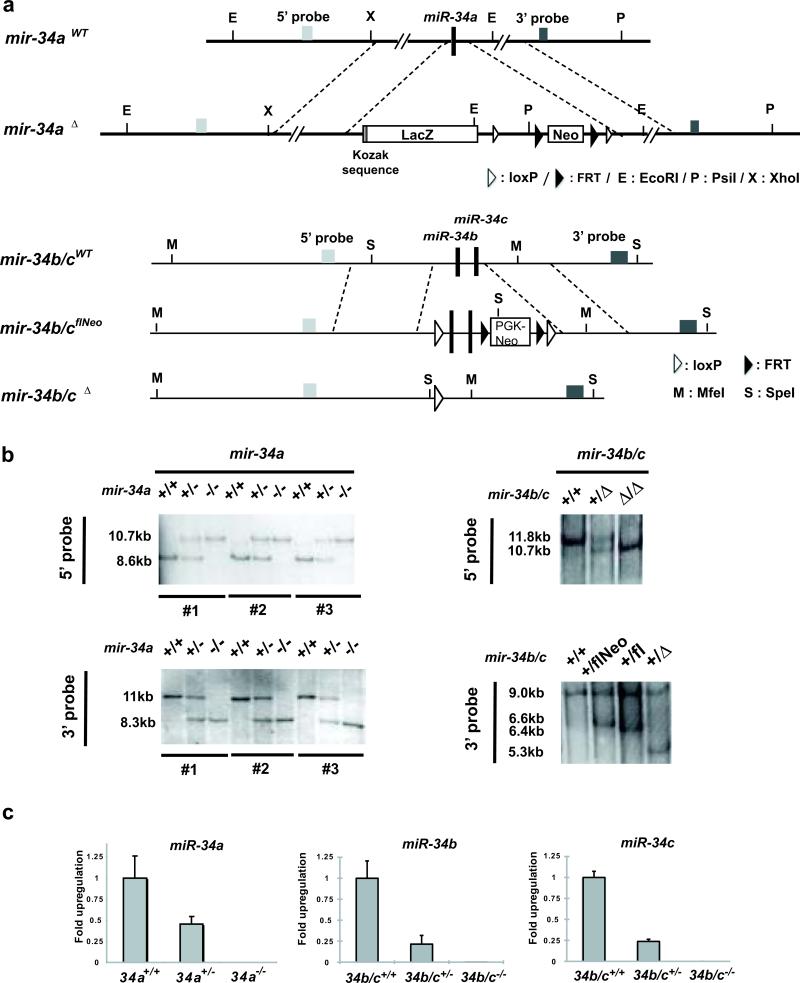
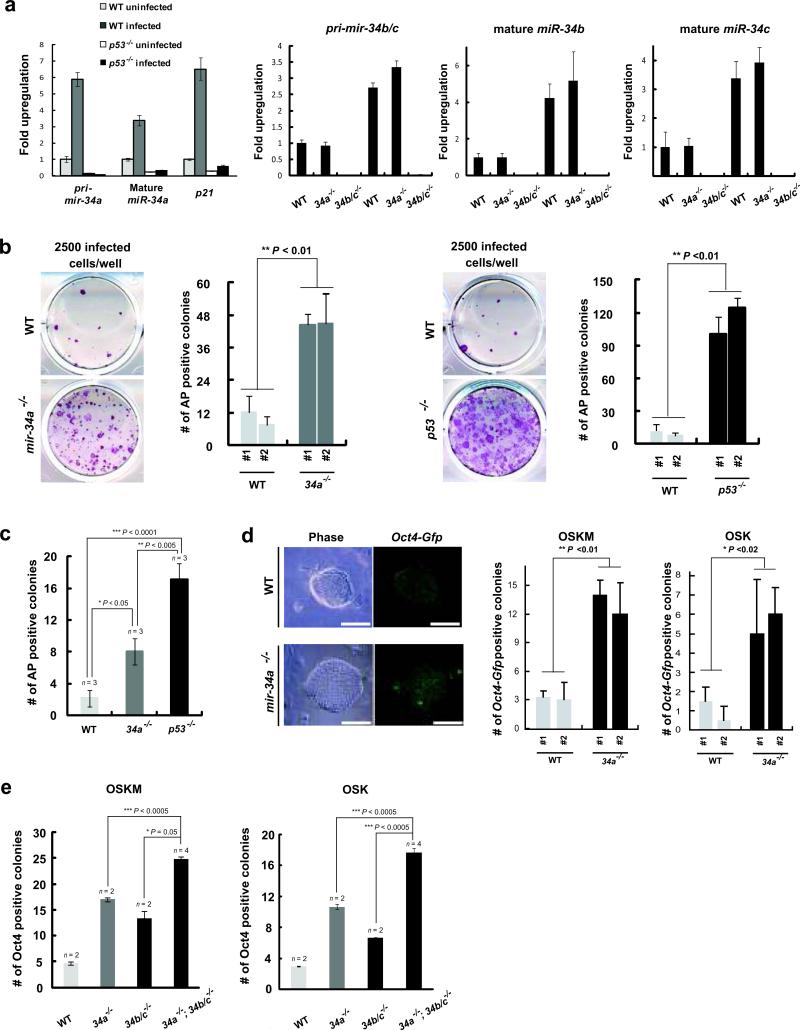
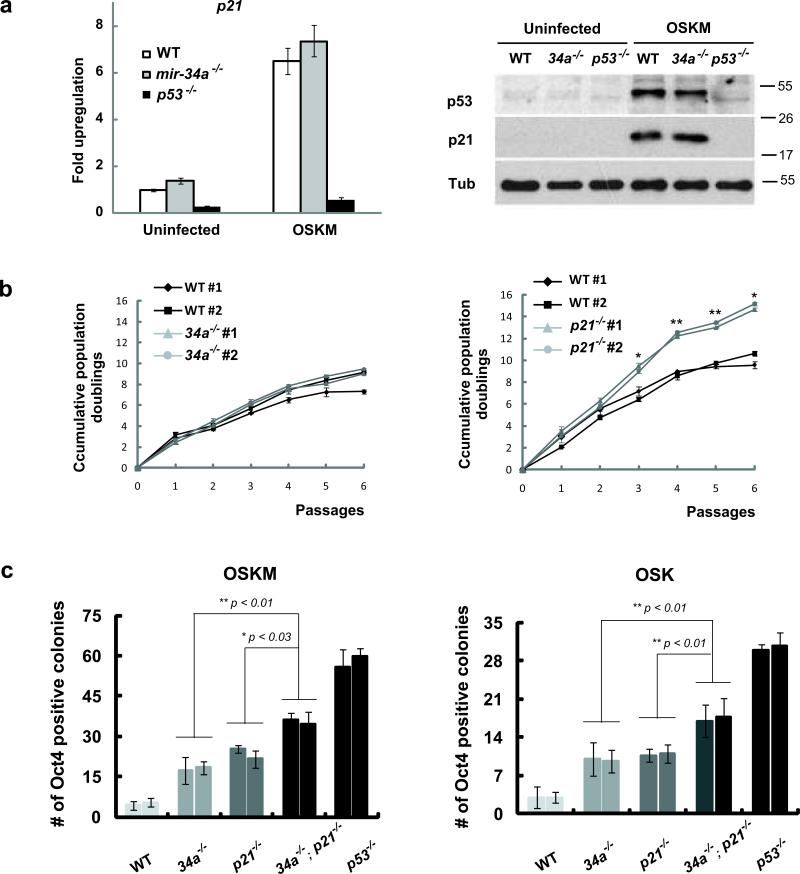
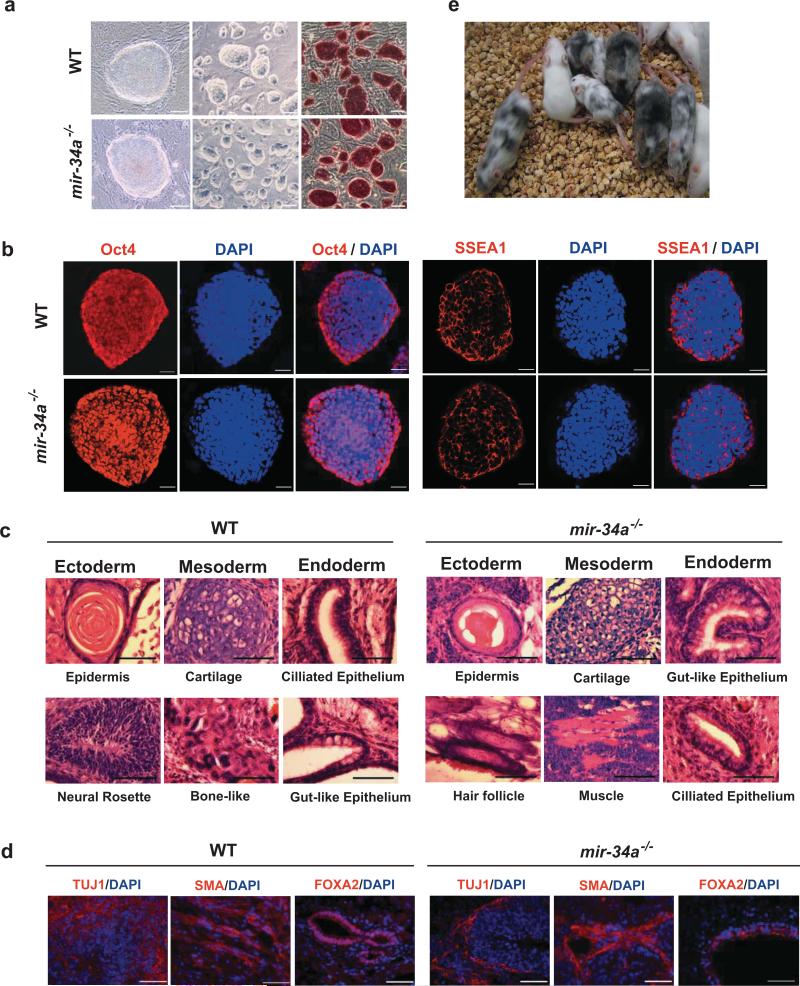
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
