

Phosphoinositide binding differentially regulates NHE1 Na+/H+ exchanger-dependent proximal tubule cell survival
- PMID: 22020933
- PMCID: PMC3234923
- DOI: 10.1074/jbc.M110.212845
Phosphoinositide binding differentially regulates NHE1 Na+/H+ exchanger-dependent proximal tubule cell survival
Abstract
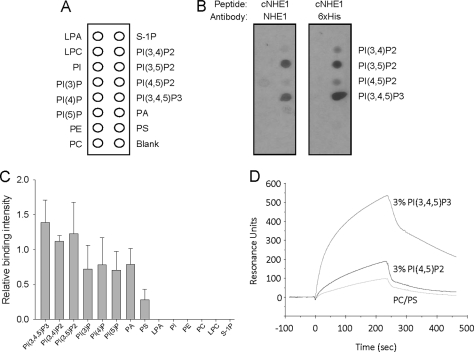
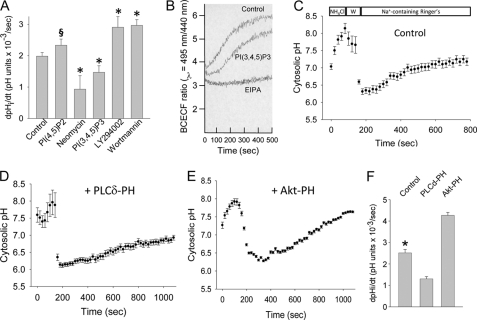
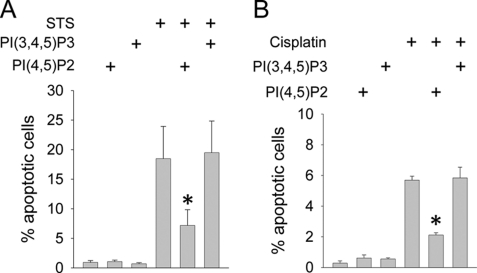
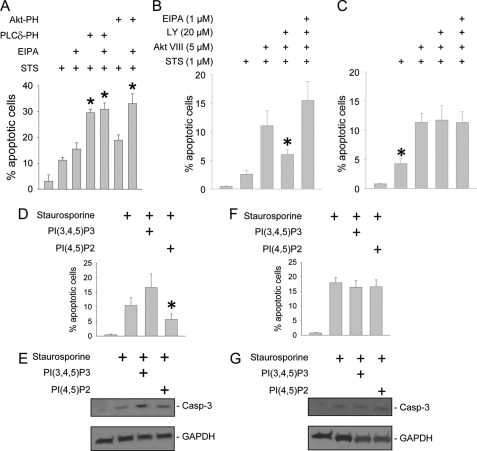
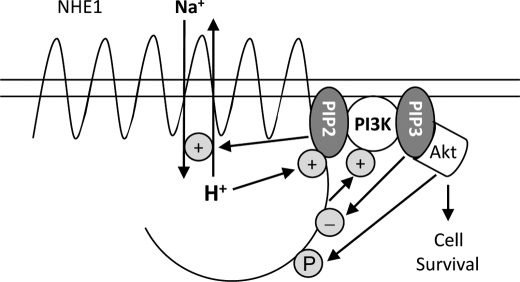
Tubular atrophy predicts chronic kidney disease progression, and is caused by proximal tubular epithelial cellcaused by proximal tubular epithelial cell (PTC) apoptosis. The normally quiescent Na(+)/H(+) exchanger-1 (NHE1) defends against PTC apoptosis, and is regulated by PI(4,5)P(2) binding. Because of the vast array of plasma membrane lipids, we hypothesized that NHE1-mediated cell survival is dynamically regulated by multiple anionic inner leaflet phospholipids. In membrane overlay and surface plasmon resonance assays, the NHE1 C terminus bound phospholipids with low affinity and according to valence (PIP(3) > PIP(2) > PIP = PA > PS). NHE1-phosphoinositide binding was enhanced by acidic pH, and abolished by NHE1 Arg/Lys to Ala mutations within two juxtamembrane domains, consistent with electrostatic interactions. PI(4,5)P(2)-incorporated vesicles were distributed to apical and lateral PTC domains, increased NHE1-regulated Na(+)/H(+) exchange, and blunted apoptosis, whereas NHE1 activity was decreased in cells enriched with PI(3,4,5)P(3), which localized to basolateral membranes. Divergent PI(4,5)P(2) and PI(3,4,5)P(3) effects on NHE1-dependent Na(+)/H(+) exchange and apoptosis were confirmed by selective phosphoinositide sequestration with pleckstrin homology domain-containing phospholipase Cδ and Akt peptides, PI 3-kinase, and Akt inhibition in wild-type and NHE1-null PTCs. The results reveal an on-off switch model, whereby NHE1 toggles between weak interactions with PI(4,5)P(2) and PI(3,4,5)P(3). In response to apoptotic stress, NHE1 is stimulated by PI(4,5)P(2), which leads to PI 3-kinase activation, and PI(4,5)P(2) phosphorylation. The resulting PI(3,4,5)P(3) dually stimulates sustained, downstream Akt survival signaling, and dampens NHE1 activity through competitive inhibition and depletion of PI(4,5)P(2).
Figures



References
-
- Coresh J., Selvin E., Stevens L. A., Manzi J., Kusek J. W., Eggers P., Van Lente F., Levey A. S. (2007) JAMA 298, 2038–2047 - PubMed
-
- Collins A. J., Foley R. N., Herzog C., Chavers B. M., Gilbertson D., Ishani A., Kasiske B. L., Liu J., Mau L. W., McBean M., Murray A., St P. W., Guo H., Li Q., Li S., Li S., Peng Y., Qiu Y., Roberts T., Skeans M., Snyder J., Solid C., Wang C., Weinhandl E., Zaun D., Arko C., Chen S. C., Dalleska F., Daniels F., Dunning S., Ebben J., Frazier E., Hanzlik C., Johnson R., Sheets D., Wang X., Forrest B., Constantini E., Everson S., Eggers P. W., Agodoa L. (2010) Am. J. Kidney Dis. 55, S1-S7 - PMC - PubMed
-
- Risdon R. A., Sloper J. C., De Wardener H. E. (1968) Lancet 2, 363–366 - PubMed
-
- Schainuck L. I., Striker G. E., Cutler R. E., Benditt E. P. (1970) Hum. Pathol. 1, 631–641 - PubMed
-
- Bohle A., Muller G. A., Wehrmann M., Mackensen-Haen S., Xiao J. C. (1996) Kidney Int. 54, S2–S9 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
