

Three structure-selective endonucleases are essential in the absence of BLM helicase in Drosophila
- PMID: 22022278
- PMCID: PMC3192830
- DOI: 10.1371/journal.pgen.1002315
Three structure-selective endonucleases are essential in the absence of BLM helicase in Drosophila
Abstract
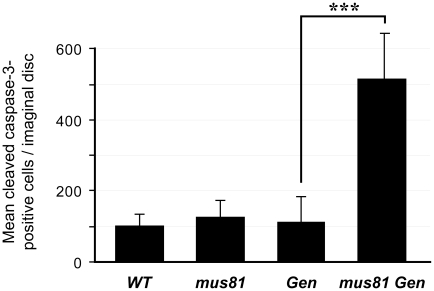
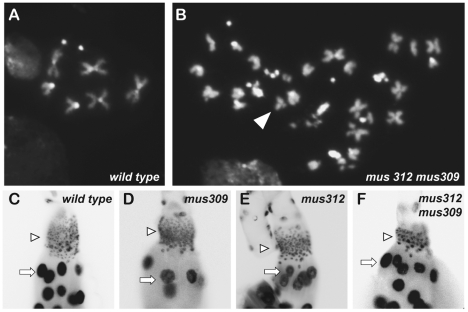
DNA repair mechanisms in mitotically proliferating cells avoid generating crossovers, which can contribute to genome instability. Most models for the production of crossovers involve an intermediate with one or more four-stranded Holliday junctions (HJs), which are resolved into duplex molecules through cleavage by specialized endonucleases. In vitro studies have implicated three nuclear enzymes in HJ resolution: MUS81-EME1/Mms4, GEN1/Yen1, and SLX4-SLX1. The Bloom syndrome helicase, BLM, plays key roles in preventing mitotic crossover, either by blocking the formation of HJ intermediates or by removing HJs without cleavage. Saccharomyces cerevisiae mutants that lack Sgs1 (the BLM ortholog) and either Mus81-Mms4 or Slx4-Slx1 are inviable, but mutants that lack Sgs1 and Yen1 are viable. The current view is that Yen1 serves primarily as a backup to Mus81-Mms4. Previous studies with Drosophila melanogaster showed that, as in yeast, loss of both DmBLM and MUS81 or MUS312 (the ortholog of SLX4) is lethal. We have now recovered and analyzed mutations in Drosophila Gen. As in yeast, there is some redundancy between Gen and mus81; however, in contrast to the case in yeast, GEN plays a more predominant role in responding to DNA damage than MUS81-MMS4. Furthermore, loss of DmBLM and GEN leads to lethality early in development. We present a comparison of phenotypes occurring in double mutants that lack DmBLM and either MUS81, GEN, or MUS312, including chromosome instability and deficiencies in cell proliferation. Our studies of synthetic lethality provide insights into the multiple functions of DmBLM and how various endonucleases may function when DmBLM is absent.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Szostak JW, Orr-Weaver TL, Rothstein RJ, Stahl FW. The double-strand-break repair model for recombination. Cell. 1983;33:25–35. - PubMed
-
- Schwacha A, Kleckner N. Identification of double Holliday junctions as intermediates in meiotic recombination. Cell. 1995;83:783–791. - PubMed
-
- German J. Bloom syndrome: a mendelian prototype of somatic mutational disease. Medicine. 1993;72:393–406. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
