

Cortical regions recruited for complex active-learning strategies and action planning exhibit rapid reactivation during memory retrieval
- PMID: 22023912
- PMCID: PMC3223278
- DOI: 10.1016/j.neuropsychologia.2011.10.012
Cortical regions recruited for complex active-learning strategies and action planning exhibit rapid reactivation during memory retrieval
Abstract
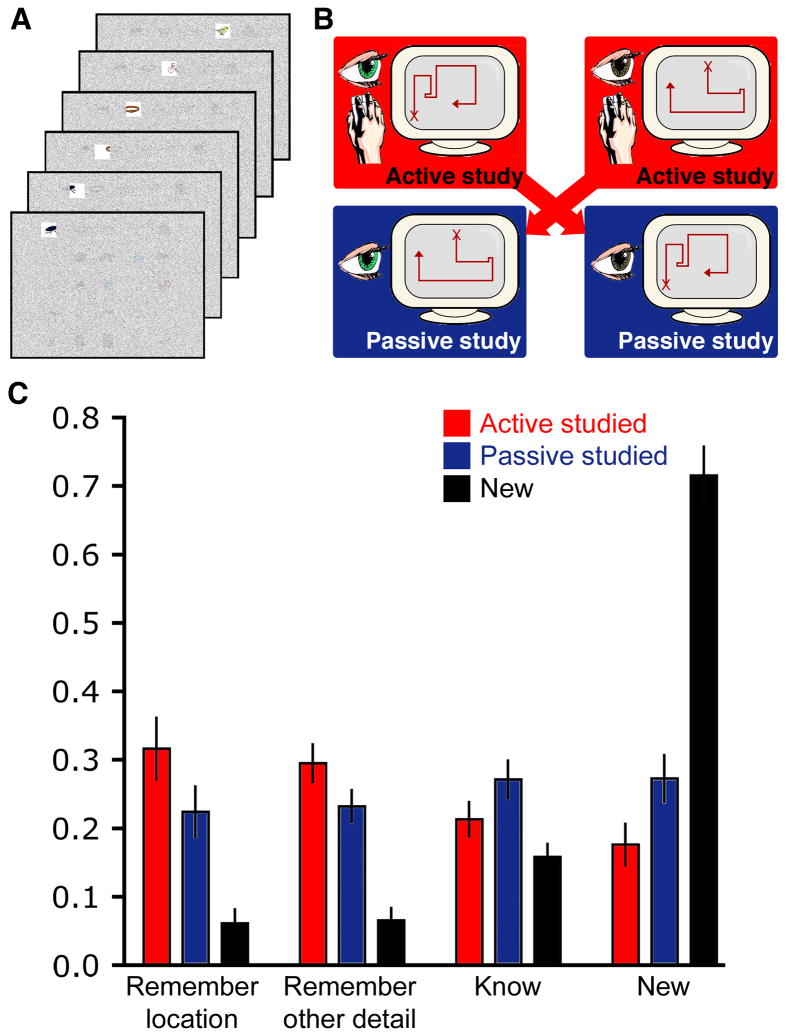
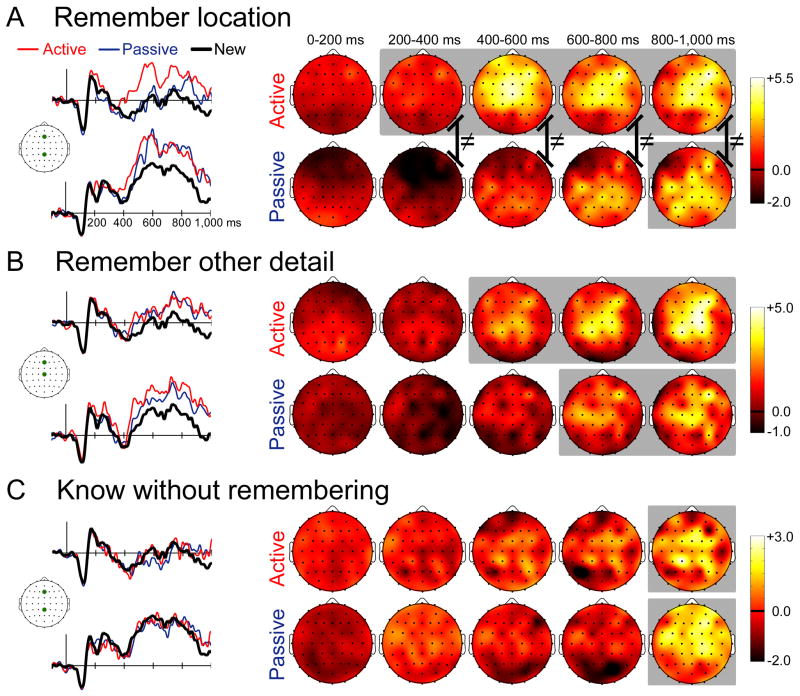
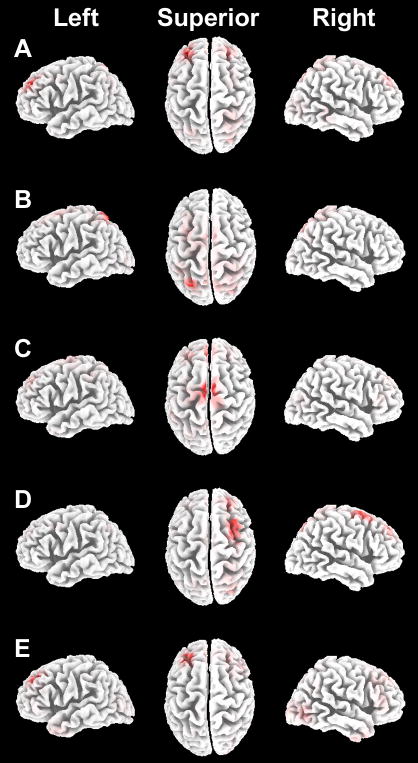
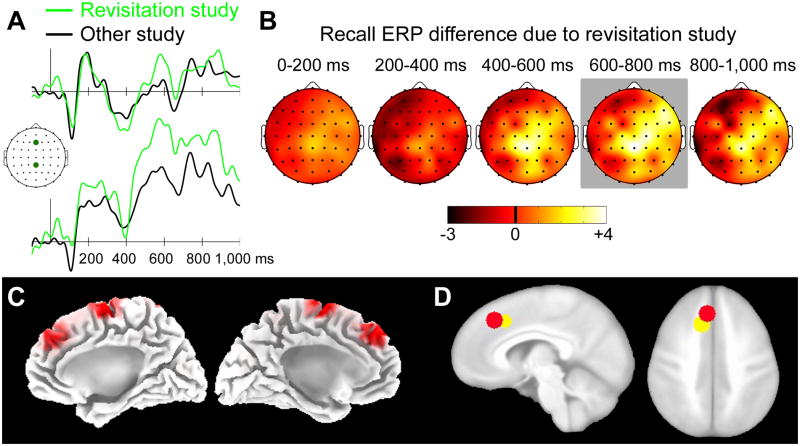
Memory retrieval can involve activity in the same sensory cortical regions involved in perception of the original event, and this neural "reactivation" has been suggested as an important mechanism of memory retrieval. However, it is still unclear if fragments of experience other than sensory information are retained and later reactivated during retrieval. For example, learning in non-laboratory settings generally involves active exploration of memoranda, thus requiring the generation of action plans for behavior and the use of strategies deployed to improve subsequent memory performance. Is information pertaining to action planning and strategic processing retained and reactivated during retrieval? To address this question, we compared ERP correlates of memory retrieval for objects that had been studied in an active manner involving action planning and strategic processing to those for objects that had been studied passively. Memory performance was superior for actively studied objects, and unique ERP retrieval correlates for these objects were identified when subjects remembered the specific spatial locations at which objects were studied. Early-onset frontal shifts in ERP correlates of retrieval were noted for these objects, which parallel the recruitment of frontal cortex during learning object locations previously identified using fMRI with the same paradigm. Notably, ERPs during recall for items studied with a specific viewing strategy localized to the same supplementary motor cortex region previously identified with fMRI when this strategy was implemented during study, suggesting rapid reactivation of regions directly involved in strategic action planning. Collectively, these results implicate neural populations involved in learning in important retrieval functions, even for those populations involved in strategic control and action planning. Notably, these episodic features are not generally reported during recollective experiences, suggesting that reactivation is a more general property of memory retrieval that extends beyond those fragments of perceptual information that might be needed to re-live the past.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Brain and behavioral indices of retrieval mode.Neuroimage. 2006 Aug 15;32(2):863-70. doi: 10.1016/j.neuroimage.2006.03.046. Epub 2006 May 15. Neuroimage. 2006. PMID: 16698287
-
Cortical activation patterns during long-term memory retrieval of visually or haptically encoded objects and locations.J Cogn Neurosci. 2009 Jan;21(1):58-82. doi: 10.1162/jocn.2009.21006. J Cogn Neurosci. 2009. PMID: 18476766
-
Individual differences in executive functioning modulate age effects on the ERP correlates of retrieval success.Neuropsychologia. 2010 Oct;48(12):3540-53. doi: 10.1016/j.neuropsychologia.2010.08.003. Epub 2010 Aug 13. Neuropsychologia. 2010. PMID: 20709089
-
The ghosts of brain states past: remembering reactivates the brain regions engaged during encoding.Psychol Bull. 2010 Jan;136(1):87-102. doi: 10.1037/a0017937. Psychol Bull. 2010. PMID: 20063927 Free PMC article. Review.
-
The neuropsychology of prospective memory in normal aging: a componential approach.Neuropsychologia. 2011 Jul;49(8):2147-55. doi: 10.1016/j.neuropsychologia.2010.12.029. Epub 2010 Dec 28. Neuropsychologia. 2011. PMID: 21192957 Free PMC article. Review.
Cited by
-
The effect of choice on intentional and incidental memory.Learn Mem. 2021 Nov 15;28(12):440-444. doi: 10.1101/lm.053433.121. Print 2021 Dec. Learn Mem. 2021. PMID: 34782402 Free PMC article.
-
Active retrieval facilitates across-episode binding by modulating the content of memory.Neuropsychologia. 2014 Oct;63:154-64. doi: 10.1016/j.neuropsychologia.2014.08.024. Epub 2014 Aug 28. Neuropsychologia. 2014. PMID: 25173711 Free PMC article.
-
The interactive effect of external rewards and self-determined choice on memory.Psychol Res. 2023 Oct;87(7):2101-2110. doi: 10.1007/s00426-023-01807-x. Epub 2023 Mar 4. Psychol Res. 2023. PMID: 36869894 Free PMC article.
-
A cross-cultural comparison of intrinsic and extrinsic motivational drives for learning.Cogn Affect Behav Neurosci. 2025 Feb;25(1):25-44. doi: 10.3758/s13415-024-01228-2. Epub 2024 Oct 18. Cogn Affect Behav Neurosci. 2025. PMID: 39424760 Free PMC article.
-
Active control over exploration improves memory in toddlers.Proc Biol Sci. 2025 Jan;292(2039):20242555. doi: 10.1098/rspb.2024.2555. Epub 2025 Jan 29. Proc Biol Sci. 2025. PMID: 39876734
References
-
- Bernstein NA. On dexterity and its development. In: Latash ML, Turvey MT, editors. Dexterity and its development. Mahwah: Lawrence Erlbaum Associates; 1996.
-
- Buckner RL, Wheeler ME. The cognitive neuroscience of remembering. Nature Reviews Neuroscience. 2001;2:624–634. - PubMed
-
- Chatrian GE, Lettich E, Nelson PL. Modified nomenclature for the “10%” electrode system. Journal of Clinical Neurophysiology. 1988;5:183–186. - PubMed
-
- Cohen NJ, Eichenbaum H. Memory, amnesia, and the hippocampal system. Cambridge: MIT Press; 1993.
-
- Conway MA, Pleydell-Pearce CW. The construction of autobiographical memories in the self-memory system. Psychological Review. 2000;107:261–288. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
