

Photoinitiated electron transfer within the Paracoccus denitrificans cytochrome bc1 complex: mobility of the iron-sulfur protein is modulated by the occupant of the Q(o) site
- PMID: 22026826
- PMCID: PMC3236023
- DOI: 10.1021/bi200453r
Photoinitiated electron transfer within the Paracoccus denitrificans cytochrome bc1 complex: mobility of the iron-sulfur protein is modulated by the occupant of the Q(o) site
Abstract
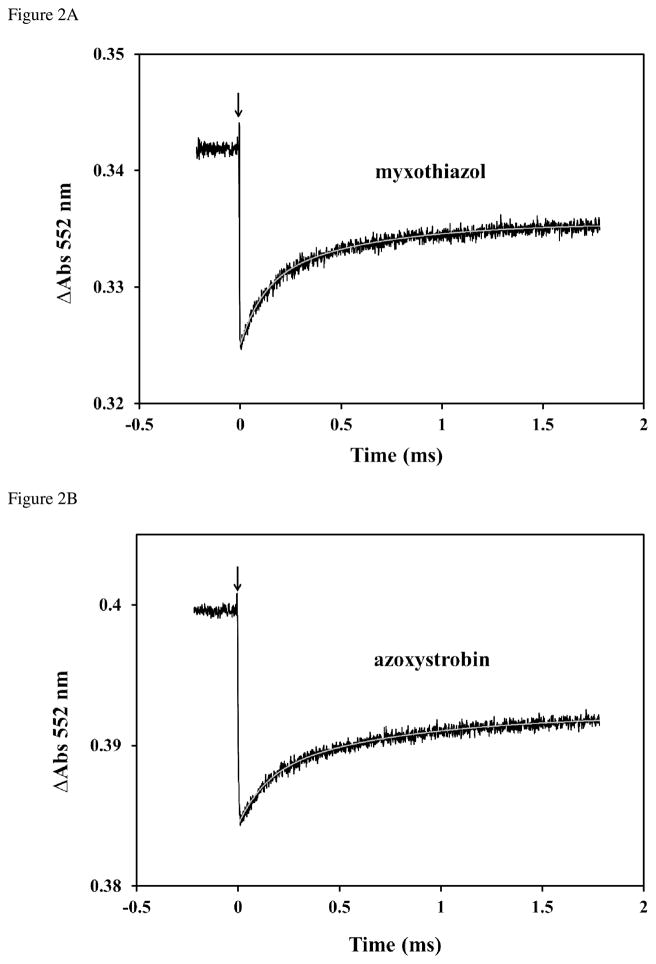
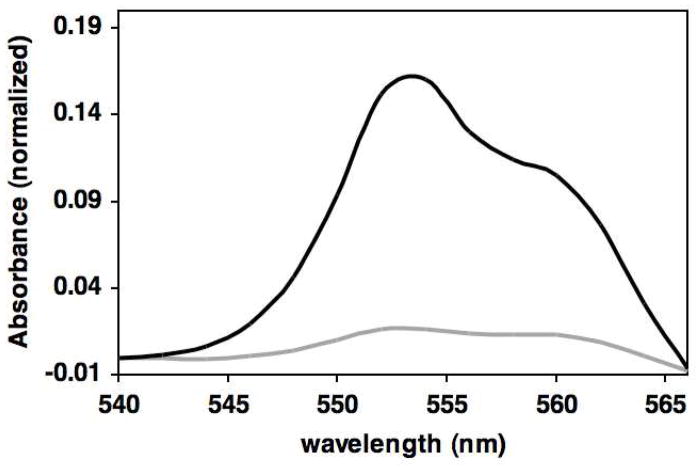
Domain rotation of the Rieske iron-sulfur protein (ISP) between the cytochrome (cyt) b and cyt c(1) redox centers plays a key role in the mechanism of the cyt bc(1) complex. Electron transfer within the cyt bc(1) complex of Paracoccus denitrificans was studied using a ruthenium dimer to rapidly photo-oxidize cyt c(1) within 1 μs and initiate the reaction. In the absence of any added quinol or inhibitor of the bc(1) complex at pH 8.0, electron transfer from reduced ISP to cyt c(1) was biphasic with rate constants of k(1f) = 6300 ± 3000 s(-1)and k(1s) = 640 ± 300 s(-1) and amplitudes of 10 ± 3% and 16 ± 4% of the total amount of cyt c(1) photooxidized. Upon addition of any of the P(m) type inhibitors MOA-stilbene, myxothiazol, or azoxystrobin to cyt bc(1) in the absence of quinol, the total amplitude increased 2-fold, consistent with a decrease in redox potential of the ISP. In addition, the relative amplitude of the fast phase increased significantly, consistent with a change in the dynamics of the ISP domain rotation. In contrast, addition of the P(f) type inhibitors JG-144 and famoxadone decreased the rate constant k(1f) by 5-10-fold and increased the amplitude over 2-fold. Addition of quinol substrate in the absence of inhibitors led to a 2-fold increase in the amplitude of the k(1f) phase. The effect of QH(2) on the kinetics of electron transfer from reduced ISP to cyt c(1) was thus similar to that of the P(m) inhibitors and very different from that of the P(f) inhibitors. The current results indicate that the species occupying the Q(o) site has a significant conformational influence on the dynamics of the ISP domain rotation.
Figures







References
-
- Trumpower BL, Gennis RB. Energy transduction by cytochrome complexes in mitochondrial and bacterial respiration: the enzymology of coupling electron transfer reactions to transmembrane proton translocation. Annu Rev Biochem. 1994;63:675–716. - PubMed
-
- Trumpower BL. The protonmotive Q cycle. Energy transduction by coupling of proton translocation to electron transfer by the cytochrome bc1 complex. J Biol Chem. 1990;265:11409–11412. - PubMed
-
- Kurowski B, Ludwig B. The genes of the Paracoccus denitrificans bc1 complex. Nucleotide sequence and homologies between bacterial and mitochondrial subunits. J Biol Chem. 1987;262:13805–13811. - PubMed
-
- Kim CH, Balny C, King TE. Role of the hinge protein in the electron transfer between cardiac cytochrome c1 and c. Equilibrium constants and kinetic probes. J Biol Chem. 1987;262:8103–8108. - PubMed
-
- Schmitt ME, Trumpower BL. Subunit 6 regulates half-of-the-sites reactivity of the dimeric cytochrome bc1 complex in Saccharomyces cerevisiae. J Biol Chem. 1990;265:17005–17011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
