

Modulation of L-α-lysophosphatidylinositol/GPR55 mitogen-activated protein kinase (MAPK) signaling by cannabinoids
- PMID: 22027819
- PMCID: PMC3249141
- DOI: 10.1074/jbc.M111.296020
Modulation of L-α-lysophosphatidylinositol/GPR55 mitogen-activated protein kinase (MAPK) signaling by cannabinoids
Abstract
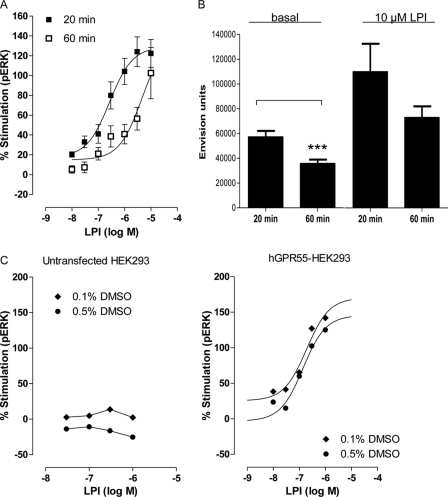
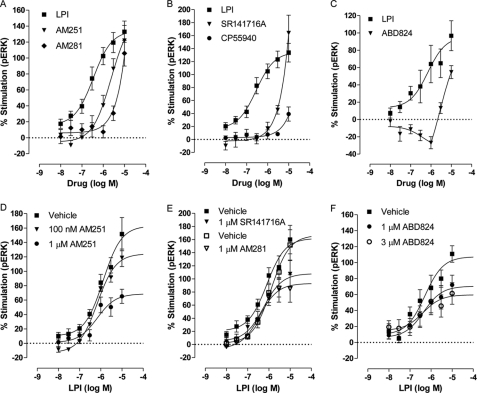
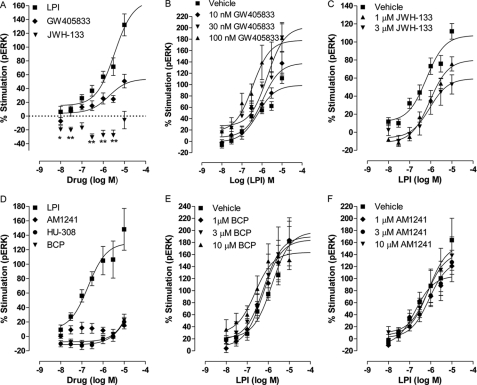
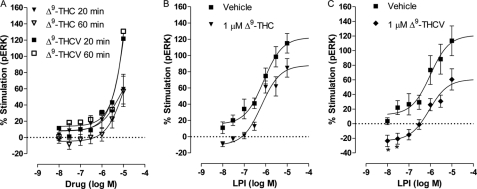
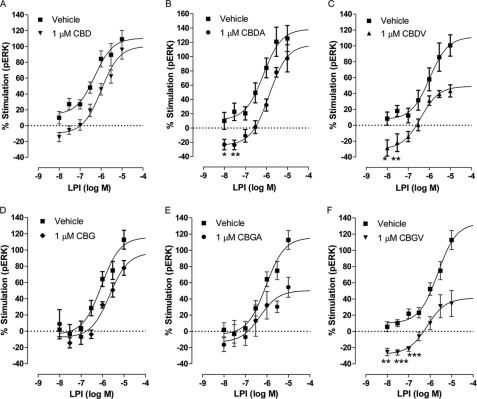
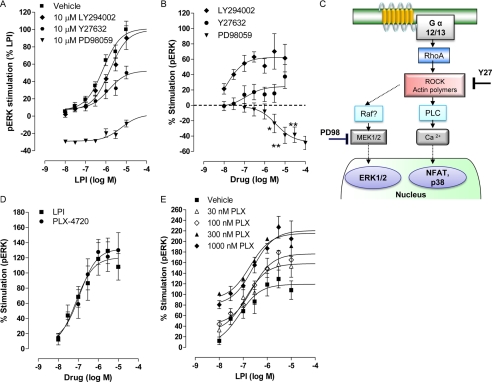
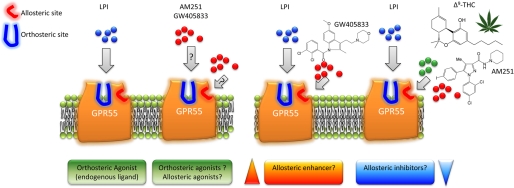
GPR55 is activated by l-α-lysophosphatidylinositol (LPI) but also by certain cannabinoids. In this study, we investigated the GPR55 pharmacology of various cannabinoids, including analogues of the CB1 receptor antagonist Rimonabant®, CB2 receptor agonists, and Cannabis sativa constituents. To test ERK1/2 phosphorylation, a primary downstream signaling pathway that conveys LPI-induced activation of GPR55, a high throughput system, was established using the AlphaScreen® SureFire® assay. Here, we show that CB1 receptor antagonists can act both as agonists alone and as inhibitors of LPI signaling under the same assay conditions. This study clarifies the controversy surrounding the GPR55-mediated actions of SR141716A; some reports indicate the compound to be an agonist and some report antagonism. In contrast, we report that the CB2 ligand GW405833 behaves as a partial agonist of GPR55 alone and enhances LPI signaling. GPR55 has been implicated in pain transmission, and thus our results suggest that this receptor may be responsible for some of the antinociceptive actions of certain CB2 receptor ligands. The phytocannabinoids Δ9-tetrahydrocannabivarin, cannabidivarin, and cannabigerovarin are also potent inhibitors of LPI. These Cannabis sativa constituents may represent novel therapeutics targeting GPR55.
Figures
References
-
- Staton P. C., Hatcher J. P., Walker D. J., Morrison A. D., Shapland E. M., Hughes J. P., Chong E., Mander P. K., Green P. J., Billinton A., Fulleylove M., Lancaster H. C., Smith J. C., Bailey L. T., Wise A., Brown A. J., Richardson J. C., Chessell I. P. (2008) Pain 139, 225–236 - PubMed
-
- Henstridge C. M., Balenga N. A., Ford L. A., Ross R. A., Waldhoer M., Irving A. J. (2009) FASEB J. 23, 183–193 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
