

Regulation of insulin-like growth factor signaling by Yap governs cardiomyocyte proliferation and embryonic heart size
- PMID: 22028467
- PMCID: PMC3440872
- DOI: 10.1126/scisignal.2002278
Regulation of insulin-like growth factor signaling by Yap governs cardiomyocyte proliferation and embryonic heart size
Abstract
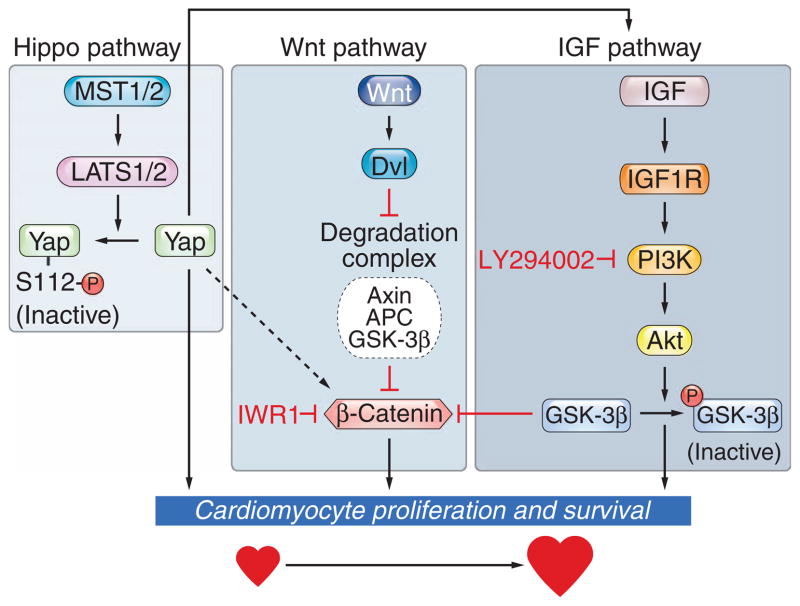
The Hippo signaling pathway regulates growth of the heart and other tissues. Hippo pathway kinases influence the activity of various targets, including the transcriptional coactivator Yap, but the specific role of Yap in heart growth has not been investigated. We show that Yap is necessary and sufficient for embryonic cardiac growth in mice. Deletion of Yap in the embryonic mouse heart impeded cardiomyocyte proliferation, causing myocardial hypoplasia and lethality at embryonic stage 10.5. Conversely, forced expression of a constitutively active form of Yap in the embryonic heart increased cardiomyocyte number and heart size. Yap activated the insulin-like growth factor (IGF) signaling pathway in cardiomyocytes, resulting in inactivation of glycogen synthase kinase 3β, which led to increased abundance of β-catenin, a positive regulator of cardiac growth. Our results point to Yap as a critical downstream effector of the Hippo pathway in the control of cardiomyocyte proliferation and a nexus for coupling the IGF, Wnt, and Hippo signaling pathways with the developmental program for heart growth.
Conflict of interest statement
Figures



References
-
- Vincent SD, Buckingham ME. How to make a heart: The origin and regulation of cardiac progenitor cells. Curr Top Dev Biol. 2010;90:1–41. - PubMed
-
- Huang J, Wu S, Barrera J, Matthews K, Pan D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila homolog of YAP. Cell. 2005;122:421–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
