

Replication timing: a fingerprint for cell identity and pluripotency
- PMID: 22028635
- PMCID: PMC3197641
- DOI: 10.1371/journal.pcbi.1002225
Replication timing: a fingerprint for cell identity and pluripotency
Abstract
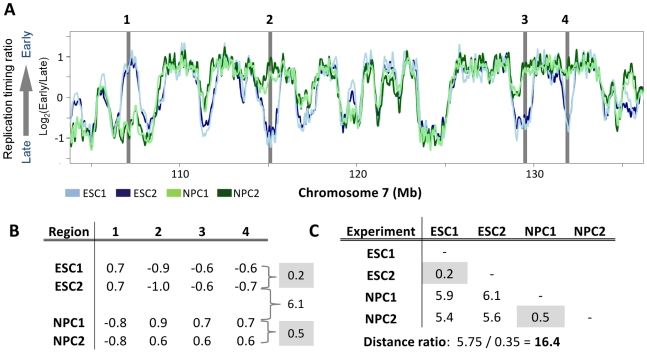
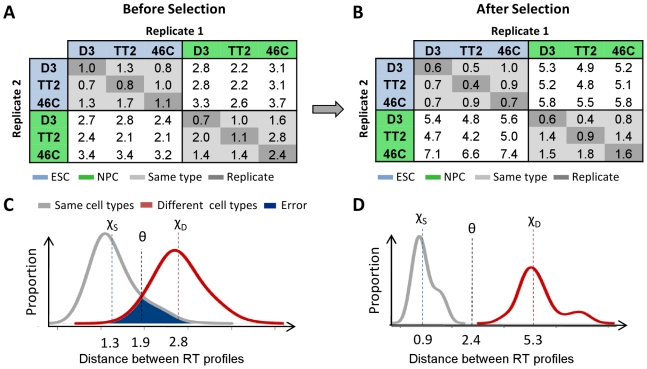
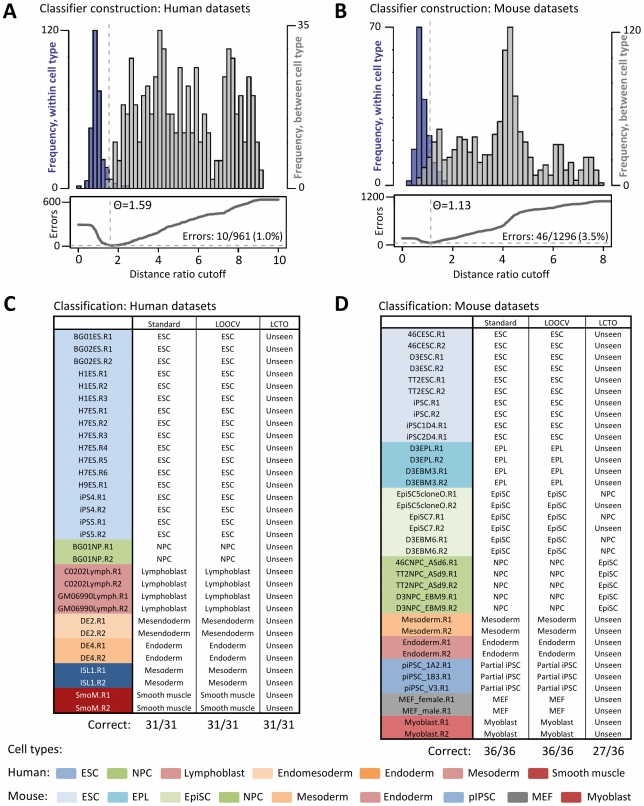
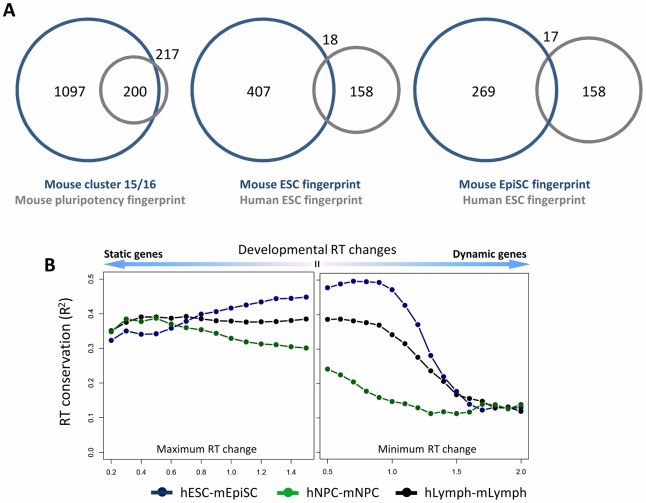
Many types of epigenetic profiling have been used to classify stem cells, stages of cellular differentiation, and cancer subtypes. Existing methods focus on local chromatin features such as DNA methylation and histone modifications that require extensive analysis for genome-wide coverage. Replication timing has emerged as a highly stable cell type-specific epigenetic feature that is regulated at the megabase-level and is easily and comprehensively analyzed genome-wide. Here, we describe a cell classification method using 67 individual replication profiles from 34 mouse and human cell lines and stem cell-derived tissues, including new data for mesendoderm, definitive endoderm, mesoderm and smooth muscle. Using a Monte-Carlo approach for selecting features of replication profiles conserved in each cell type, we identify "replication timing fingerprints" unique to each cell type and apply a k nearest neighbor approach to predict known and unknown cell types. Our method correctly classifies 67/67 independent replication-timing profiles, including those derived from closely related intermediate stages. We also apply this method to derive fingerprints for pluripotency in human and mouse cells. Interestingly, the mouse pluripotency fingerprint overlaps almost completely with previously identified genomic segments that switch from early to late replication as pluripotency is lost. Thereafter, replication timing and transcription within these regions become difficult to reprogram back to pluripotency, suggesting these regions highlight an epigenetic barrier to reprogramming. In addition, the major histone cluster Hist1 consistently becomes later replicating in committed cell types, and several histone H1 genes in this cluster are downregulated during differentiation, suggesting a possible instrument for the chromatin compaction observed during differentiation. Finally, we demonstrate that unknown samples can be classified independently using site-specific PCR against fingerprint regions. In sum, replication fingerprints provide a comprehensive means for cell characterization and are a promising tool for identifying regions with cell type-specific organization.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Woodfine K, Fiegler H, Beare DM, Collins JE, McCann OT, et al. Replication timing of the human genome. Hum Mol Genet. 2004;13:191–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
