

Identification of widespread ultra-edited human RNAs
- PMID: 22028664
- PMCID: PMC3197674
- DOI: 10.1371/journal.pgen.1002317
Identification of widespread ultra-edited human RNAs
Abstract
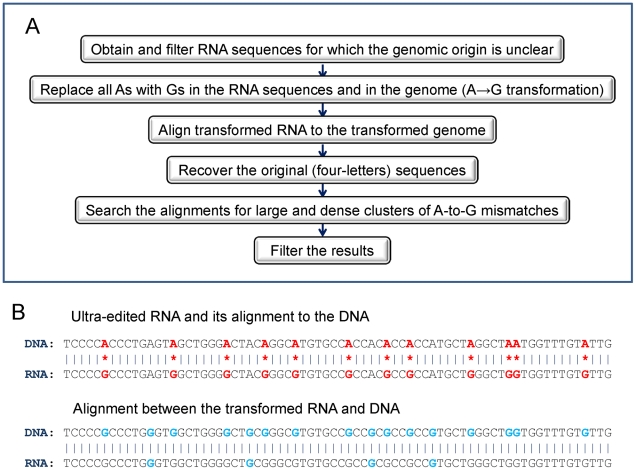
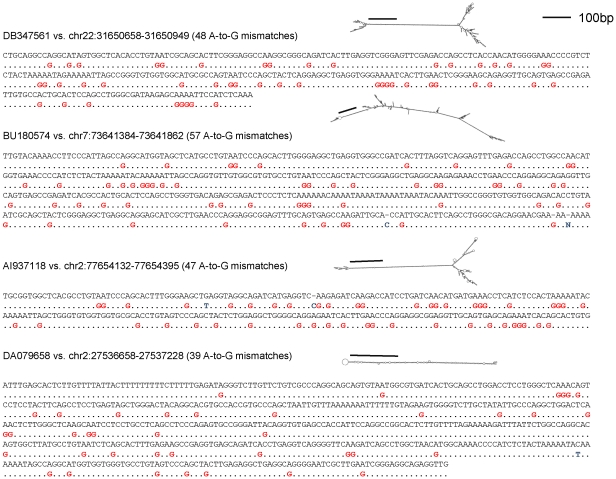
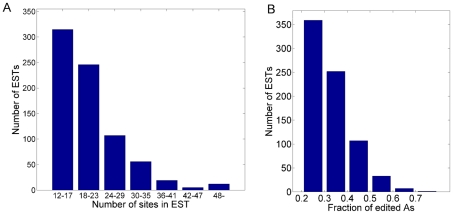
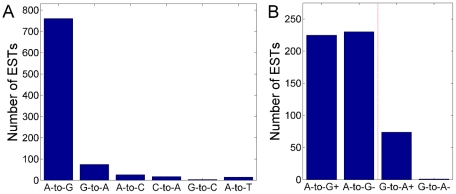
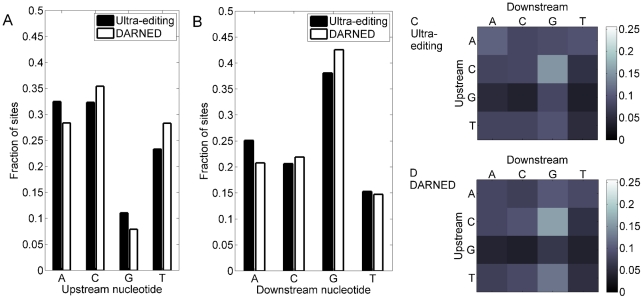
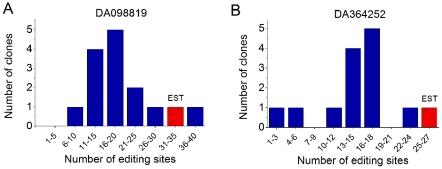
Adenosine-to-inosine modification of RNA molecules (A-to-I RNA editing) is an important mechanism that increases transciptome diversity. It occurs when a genomically encoded adenosine (A) is converted to an inosine (I) by ADAR proteins. Sequencing reactions read inosine as guanosine (G); therefore, current methods to detect A-to-I editing sites align RNA sequences to their corresponding DNA regions and identify A-to-G mismatches. However, such methods perform poorly on RNAs that underwent extensive editing ("ultra"-editing), as the large number of mismatches obscures the genomic origin of these RNAs. Therefore, only a few anecdotal ultra-edited RNAs have been discovered so far. Here we introduce and apply a novel computational method to identify ultra-edited RNAs. We detected 760 ESTs containing 15,646 editing sites (more than 20 sites per EST, on average), of which 13,668 are novel. Ultra-edited RNAs exhibit the known sequence motif of ADARs and tend to localize in sense strand Alu elements. Compared to sites of mild editing, ultra-editing occurs primarily in Alu-rich regions, where potential base pairing with neighboring, inverted Alus creates particularly long double-stranded RNA structures. Ultra-editing sites are underrepresented in old Alu subfamilies, tend to be non-conserved, and avoid exons, suggesting that ultra-editing is usually deleterious. A possible biological function of ultra-editing could be mediated by non-canonical splicing and cleavage of the RNA near the editing sites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
