

Human evolution and osteoporosis-related spinal fractures
- PMID: 22028933
- PMCID: PMC3197574
- DOI: 10.1371/journal.pone.0026658
Human evolution and osteoporosis-related spinal fractures
Abstract
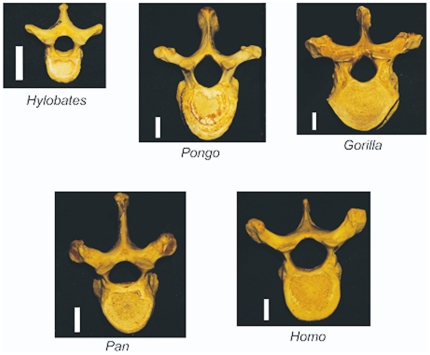
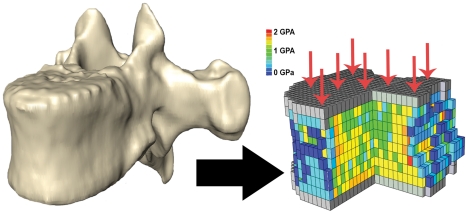
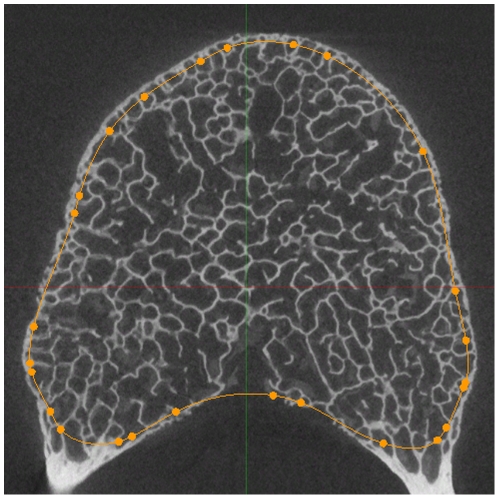
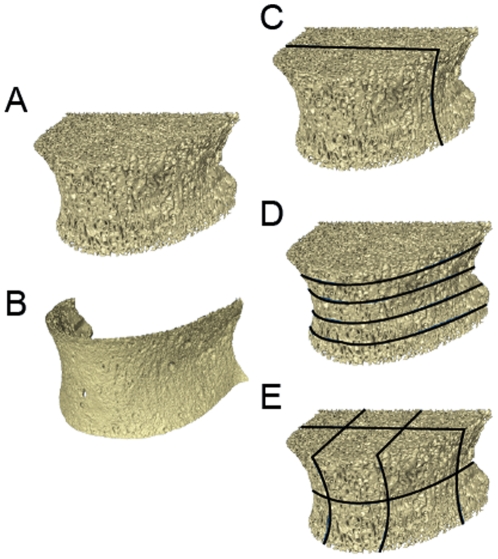
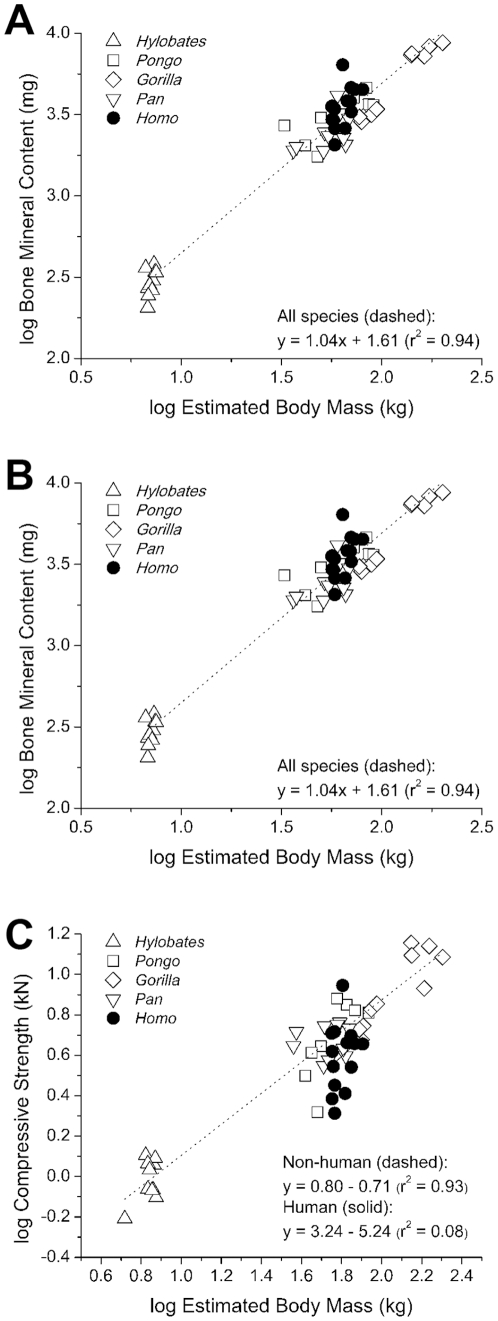
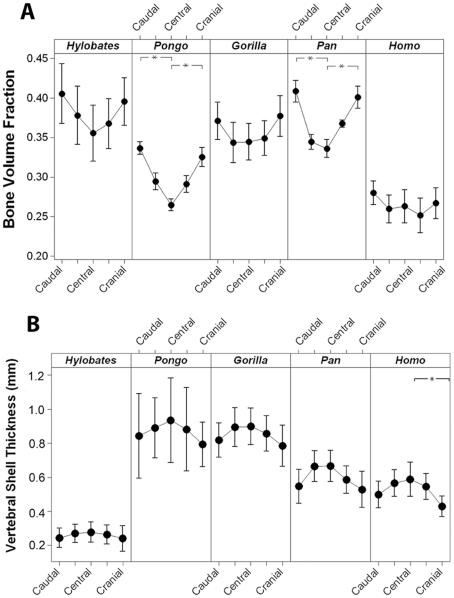
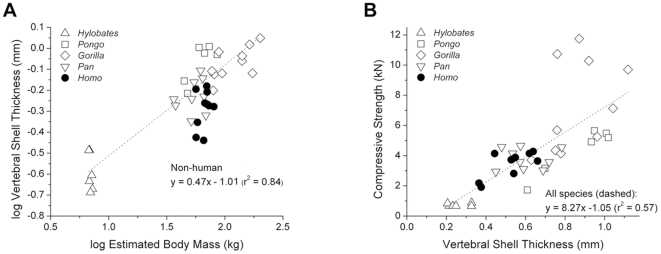
The field of evolutionary medicine examines the possibility that some diseases are the result of trade-offs made in human evolution. Spinal fractures are the most common osteoporosis-related fracture in humans, but are not observed in apes, even in cases of severe osteopenia. In humans, the development of osteoporosis is influenced by peak bone mass and strength in early adulthood as well as age-related bone loss. Here, we examine the structural differences in the vertebral bodies (the portion of the vertebra most commonly involved in osteoporosis-related fractures) between humans and apes before age-related bone loss occurs. Vertebrae from young adult humans and chimpanzees, gorillas, orangutans, and gibbons (T8 vertebrae, n = 8-14 per species, male and female, humans: 20-40 years of age) were examined to determine bone strength (using finite element models), bone morphology (external shape), and trabecular microarchitecture (micro-computed tomography). The vertebrae of young adult humans are not as strong as those from apes after accounting for body mass (p<0.01). Human vertebrae are larger in size (volume, cross-sectional area, height) than in apes with a similar body mass. Young adult human vertebrae have significantly lower trabecular bone volume fraction (0.26±0.04 in humans and 0.37±0.07 in apes, mean ± SD, p<0.01) and thinner vertebral shells than apes (after accounting for body mass, p<0.01). Since human vertebrae are more porous and weaker than those in apes in young adulthood (after accounting for bone mass), even modest amounts of age-related bone loss may lead to vertebral fracture in humans, while in apes, larger amounts of bone loss would be required before a vertebral fracture becomes likely. We present arguments that differences in vertebral bone size and shape associated with reduced bone strength in humans is linked to evolutionary adaptations associated with bipedalism.
Conflict of interest statement
Figures
References
-
- Trevathan W, Smith EO, McKenna JJ, editors. Evolutionary Medicine and Health: New Perspectives. New York: Oxford University Press; 2008. An overview of evolutionary medicine.
-
- Sambrook P, Cooper C. Osteoporosis. Lancet. 2006;367:2010–2018. - PubMed
-
- Gunji H, Hosaka K, Huffman MA, Kawanaka K, Matsumoto-Oda A, et al. Extraordinarily low bone mineral density in an old female chimpanzee (Pan troglodytes schweinfurthii) from the Mahale Mountains National Park. Primates. 2003;44:145–149. - PubMed
-
- Jurmain R. Degenerative joint disease in African great apes: an evolutionary perspective. Journal of Human Evolution. 2000;39:185–203. - PubMed
-
- Lovell NC. Washington DC: Smithsonian Institution Press; 1990. Patterns of injury and illness in great apes.224
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
