

Bmal1 in the nervous system is essential for normal adaptation of circadian locomotor activity and food intake to periodic feeding
- PMID: 22031885
- PMCID: PMC6703508
- DOI: 10.1523/JNEUROSCI.2801-11.2011
Bmal1 in the nervous system is essential for normal adaptation of circadian locomotor activity and food intake to periodic feeding
Abstract
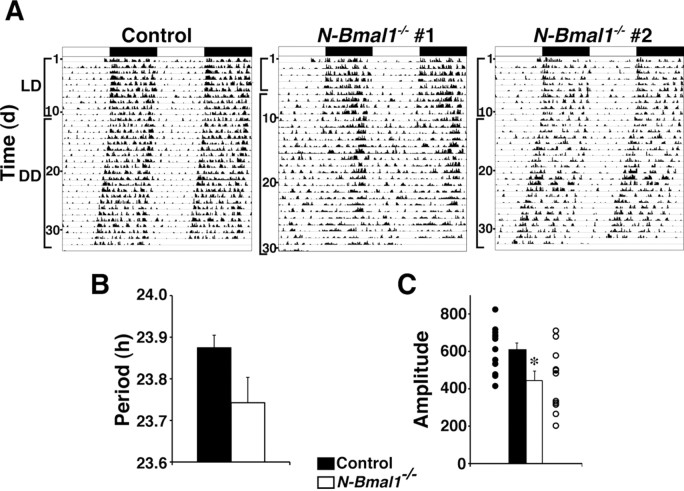
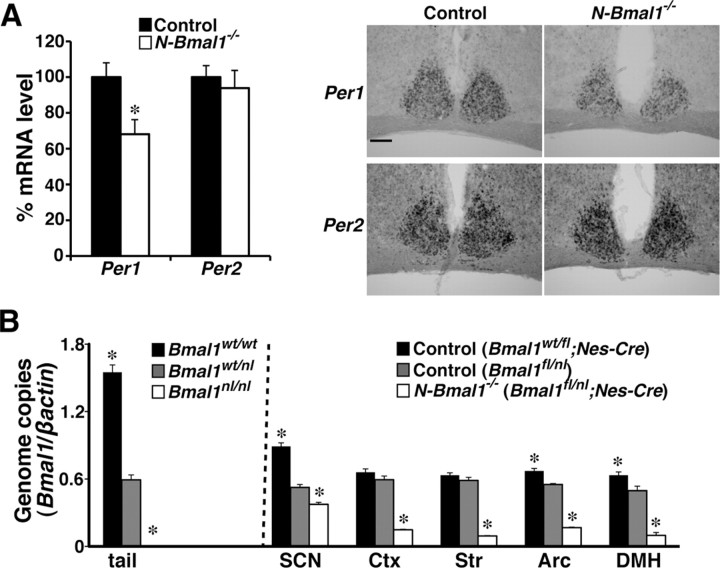
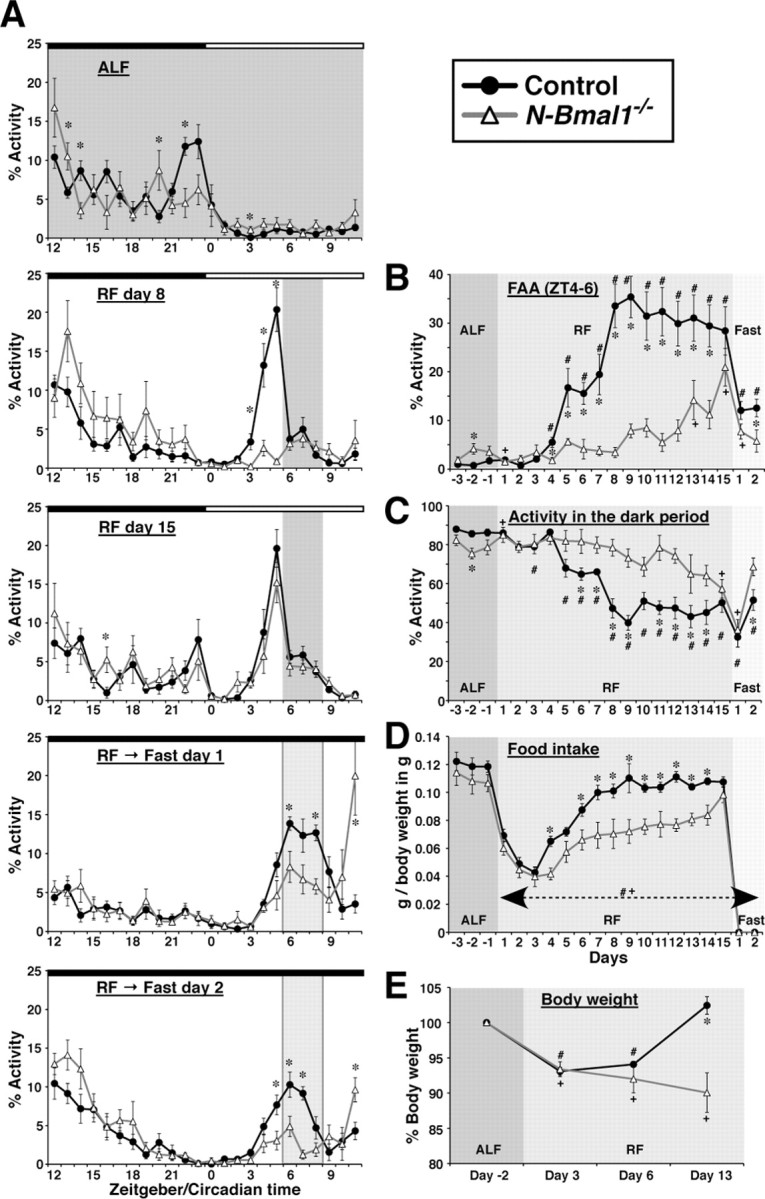
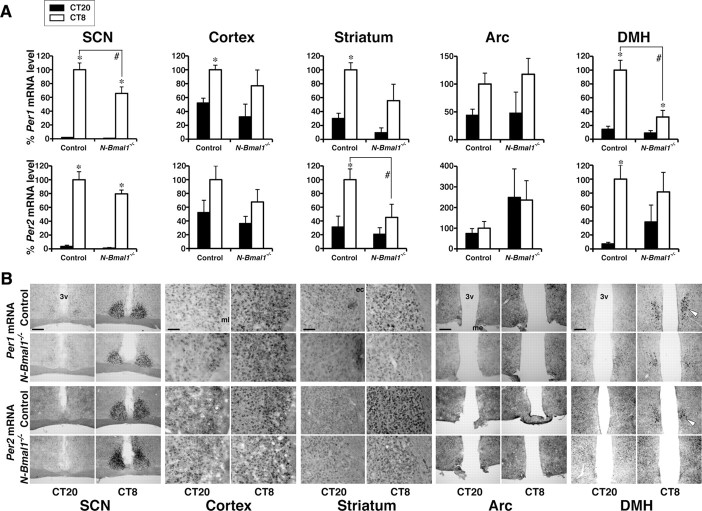
Temporal restriction of feeding can entrain circadian behavioral and physiological rhythms in mammals. These changes in biological rhythms are postulated to be brought about by a putative food-entrainable oscillator (FEO) that is independent of the suprachiasmatic nucleus (SCN). However, the anatomical substrates and molecular machinery of FEO remain elusive. We report here that mice with a nervous system-specific deletion of Bmal1, an essential clock component, had a marked deficit in entrainment of locomotor activity by periodic feeding, accompanied by reduced food intake and subsequent loss of body weight. These mice exhibited a nearly normal light-entrainable activity rhythm driven by the SCN, because deletion of the Bmal1 gene in the SCN was only partial. These findings suggest that an SCN-independent FEO in the nervous system requires Bmal1 and plays a critical role in adaptation of circadian locomotor activity and food intake to periodic feeding.
Figures
References
-
- Acosta-Galvan G, Yi CX, van der Vliet J, Jhamandas JH, Panula P, Angeles-Castellanos M, Del Carmen Basualdo M, Escobar C, Buijs RM. Interaction between hypothalamic dorsomedial nucleus and the suprachiasmatic nucleus determines intensity of food anticipatory behavior. Proc Natl Acad Sci U S A. 2011;108:5813–5818. - PMC - PubMed
-
- Bunger MK, Walisser JA, Sullivan R, Manley PA, Moran SM, Kalscheur VL, Colman RJ, Bradfield CA. Progressive arthropathy in mice with a targeted disruption of the Mop3/Bmal-1 locus. Genesis. 2005;41:122–132. - PubMed
-
- Challet E, Mendoza J, Dardente H, Pévet P. Neurogenetics of food anticipation. Eur J Neurosci. 2009;30:1676–1687. - PubMed
-
- Davidson AJ. Search for the feeding-entrainable circadian oscillator: a complex proposition. Am J Physiol Regul Integr Comp Physiol. 2006;290:R1524–R1526. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases