

Diversity and modularity of G protein-coupled receptor structures
- PMID: 22032986
- PMCID: PMC3259144
- DOI: 10.1016/j.tips.2011.09.003
Diversity and modularity of G protein-coupled receptor structures
Abstract
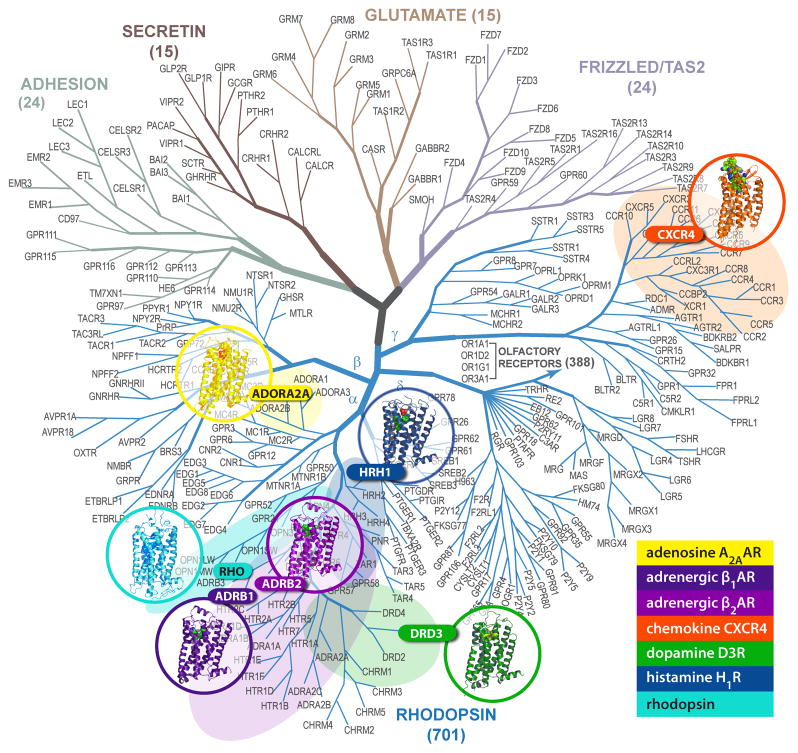
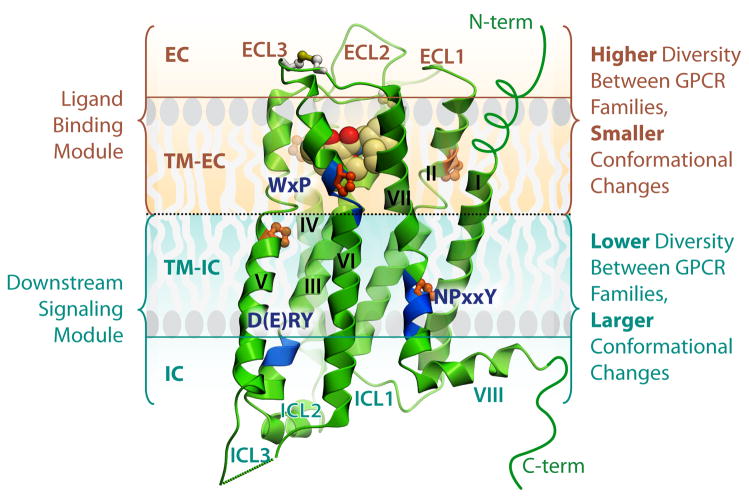
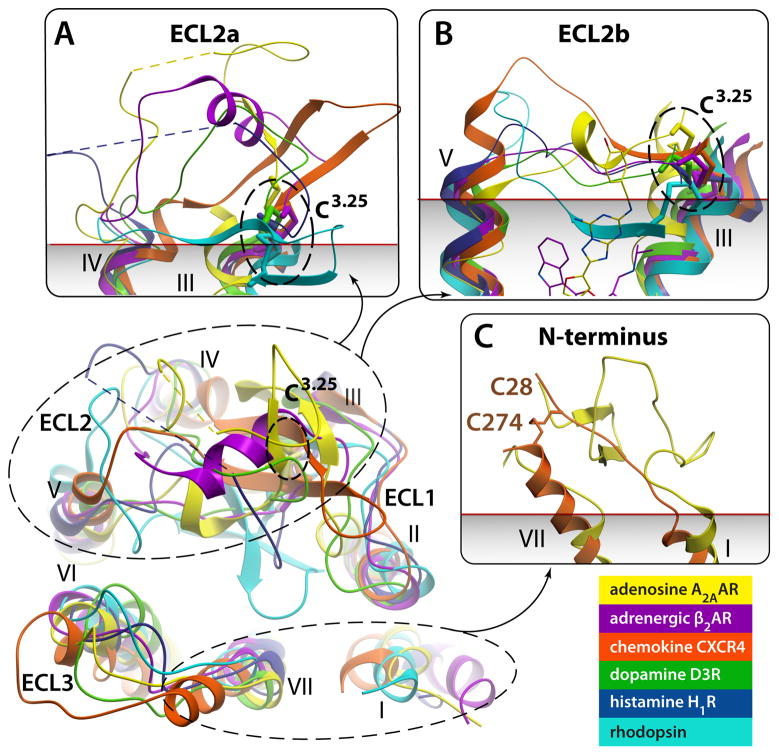
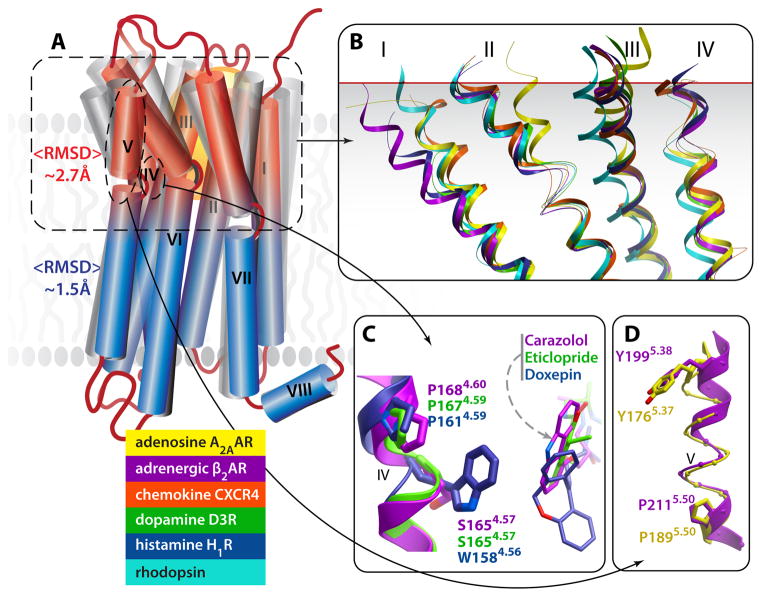
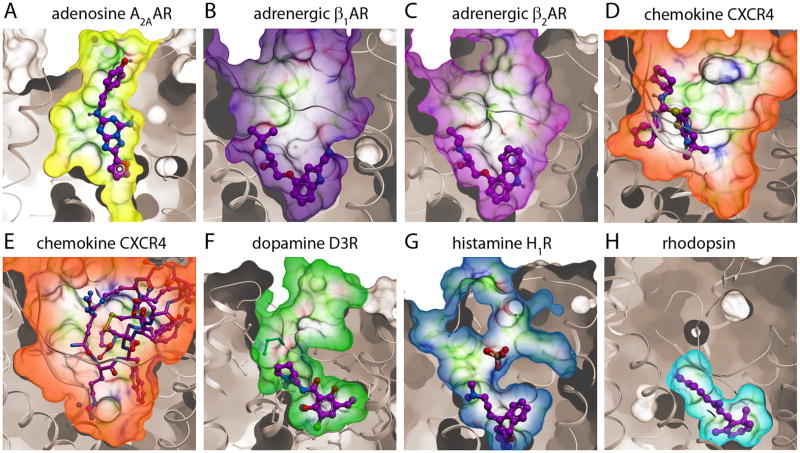
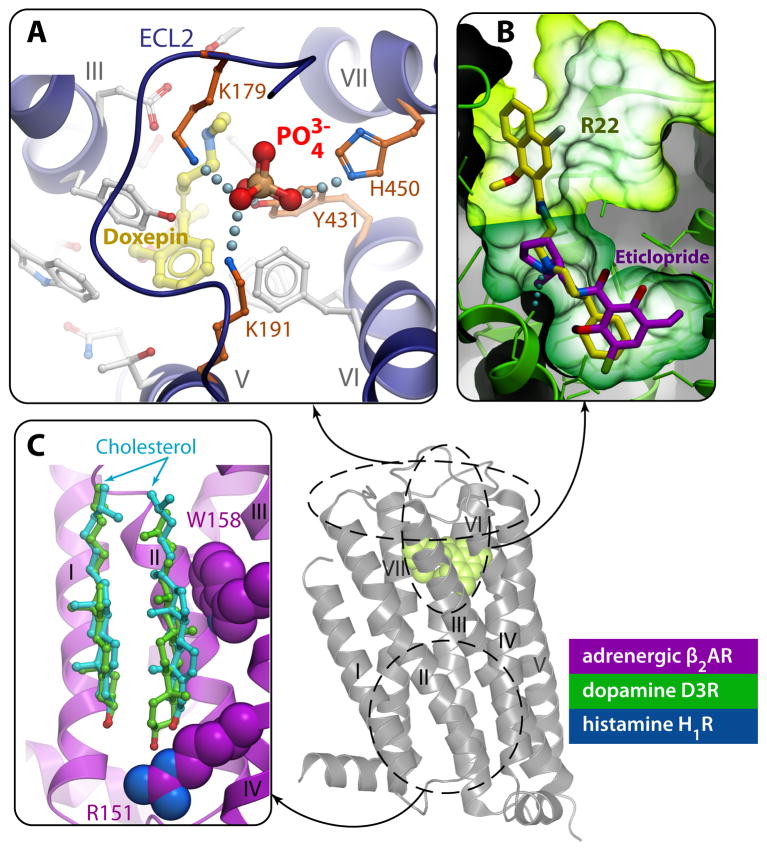
G protein-coupled receptors (GPCRs) comprise the most 'prolific' family of cell membrane proteins, which share a general mechanism of signal transduction, but greatly vary in ligand recognition and function. Crystal structures are now available for rhodopsin, adrenergic, and adenosine receptors in both inactive and activated forms, as well as for chemokine, dopamine, and histamine receptors in inactive conformations. Here we review common structural features, outline the scope of structural diversity of GPCRs at different levels of homology, and briefly discuss the impact of the structures on drug discovery. Given the current set of GPCR crystal structures, a distinct modularity is now being observed between the extracellular (ligand-binding) and intracellular (signaling) regions. The rapidly expanding repertoire of GPCR structures provides a solid framework for experimental and molecular modeling studies, and helps to chart a roadmap for comprehensive structural coverage of the whole superfamily and an understanding of GPCR biological and therapeutic mechanisms.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Nordstrom KJ, et al. Independent HHsearch, Needleman-Wunsch-based and motif analyses reveals the overall hierarchy for most of the G protein-coupled receptor families. Mol Biol Evol. 2011;28(9):2471–80. - PubMed
-
- Fredriksson R, et al. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol. 2003;63(6):1256–72. - PubMed
-
- Fredriksson R, Schioth HB. The repertoire of G-protein-coupled receptors in fully sequenced genomes. Mol Pharmacol. 2005;67(5):1414–25. - PubMed
-
- Strotmann R, et al. Evolution of GPCR: change and continuity. Mol Cell Endocrinol. 2011;331(2):170–8. - PubMed
-
- Lagerstrom MC, Schioth HB. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat Rev Drug Discov. 2008;7(4):339–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
