

Review
doi: 10.1038/nmeth.1735.
Characterizing RNA dynamics at atomic resolution using solution-state NMR spectroscopy
Affiliations
- PMID: 22036746
- PMCID: PMC3320163
- DOI: 10.1038/nmeth.1735
Item in Clipboard
Review
Characterizing RNA dynamics at atomic resolution using solution-state NMR spectroscopy
Nat Methods.
.
Abstract
Many recently discovered noncoding RNAs do not fold into a single native conformation but sample many different conformations along their free-energy landscape to carry out their biological function. Here we review solution-state NMR techniques that measure the structural, kinetic and thermodynamic characteristics of RNA motions spanning picosecond to second timescales at atomic resolution, allowing unprecedented insights into the RNA dynamic structure landscape. From these studies a basic description of the RNA dynamic structure landscape is emerging, bringing new insights into how RNA structures change to carry out their function as well as applications in RNA-targeted drug discovery and RNA bioengineering.
Figures
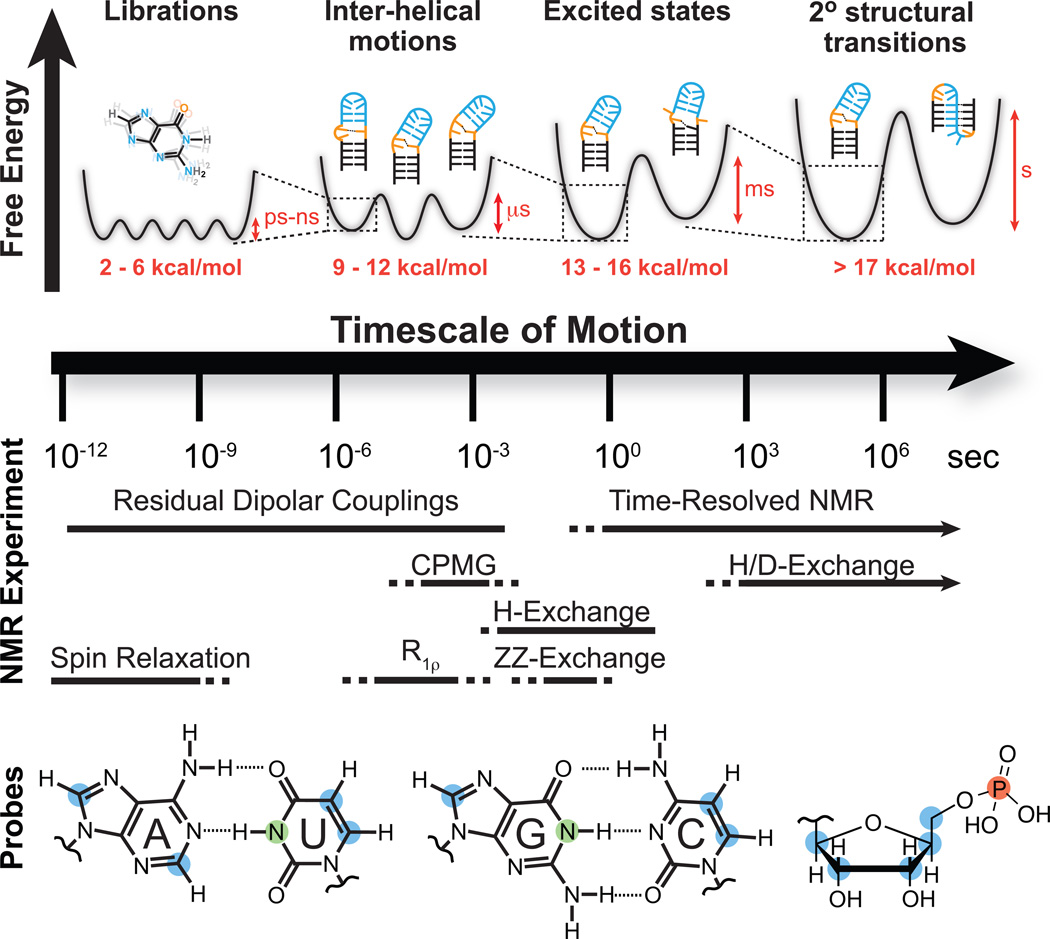
NMR techniques and site-specific probes for characterizing motional modes that carry RNA structure along various regions of the dynamic structure landscape. Dynamic Structure Landscape: Transition free energies (red, kcal/mol) corresponding to typical timescales of interconversion were estimated using transition state theory at 25 °C. NMR Experiments: Solid lines indicate the timescales at which each NMR experiment is optimally suited for, while the dotted lines indicate timescales that are difficult to probe. Probes: Nuclei most commonly used for RNA dynamics measurements: protonated carbons (blue), imino protons (green), backbone phosphorus (red).
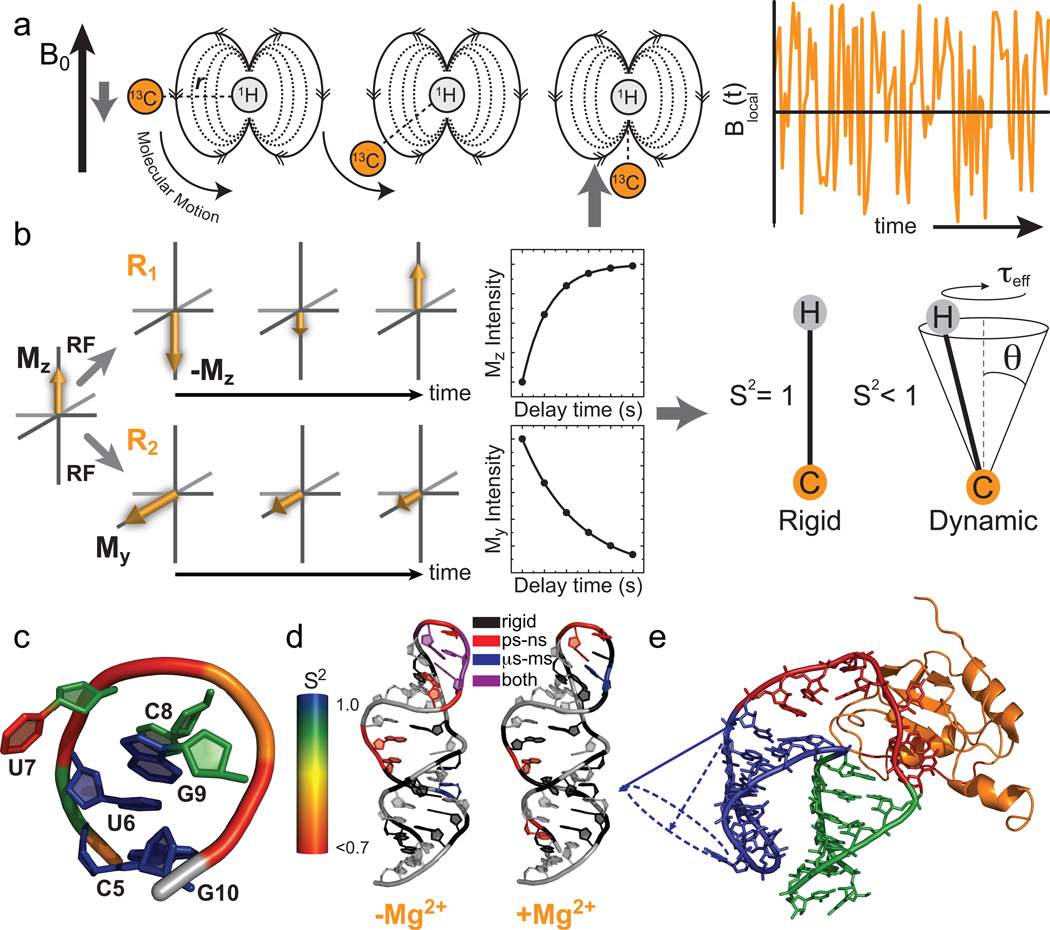
Characterization of pico- to nanosecond motions using spin relaxation. (a) Reorientation of bond vectors leads to an oscillating local field at the nucleus of interest that influences relaxation. Shown is an example involving a 13C-1H dipolar interaction. (b) Model free analysis of longitudinal (R1) and transverse (R2) spin relaxation data yield an order parameter (S2) describing the amplitude of motion and a constant (τeff) describing its timescale. (c–e) Example applications of spin relaxation in studies of RNA dynamics, (c) Site-specific UUCG tetraloop dynamics. Note, the sugar and nucleobase order parameters, were determined at 298 K while the phosphate backbone order parameters were determined at 310 K. (d) Re-distribution of motional modes in the catalytic domain 5 RNA of a group II intron upon addition of Mg2+ (Adapted from ref. with permission from Elsevier). (e) RNA inter-helical motions persist when bound to the human U1A protein (Reprinted with permission from ref. . Copyright 2007 American Chemical Society).
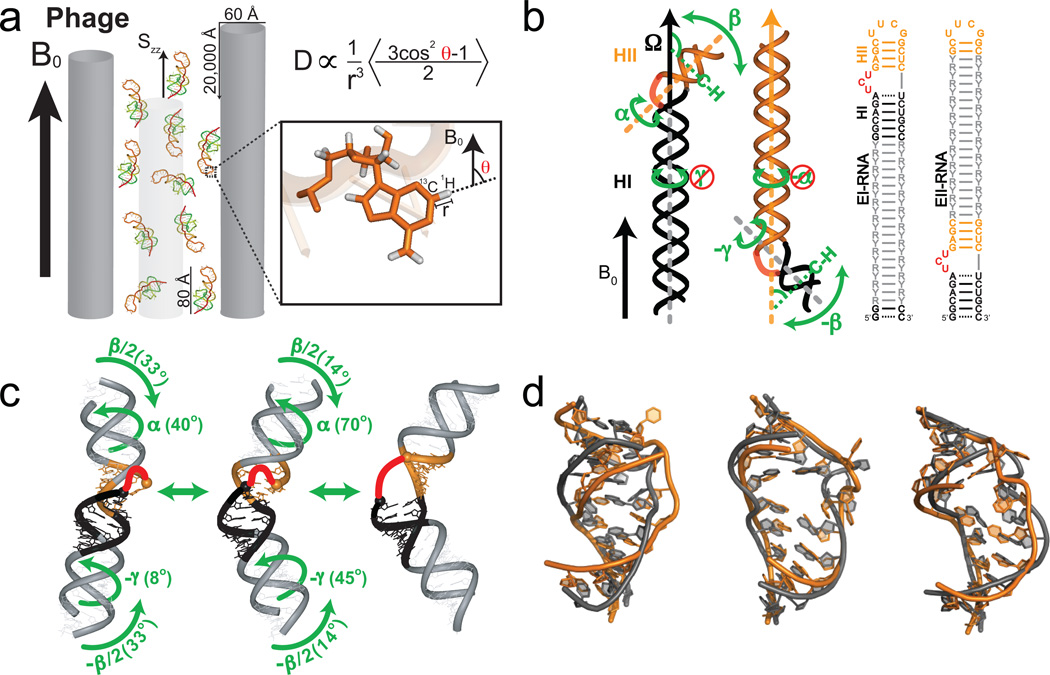
Characterizing motions over sub-millisecond timescales using residual dipolar couplings. (a) Partial alignment of RNA using Pf1 phage shown as gray rods (left) (Adapted from ref. 43). (b) Domain-elongation for decoupling internal and overall motions allows measurement of bond vector dynamics relative to the elongated helix. An isotopic labeling strategy is used to render elongation residues (R-Y) NMR invisible (adapted from ref. 52). (c) Spatially correlated inter-helical motions observed using RDCs involving correlated changes in the inter-helical twist (α and γ) and bend (β) angles (adapted from ref. 52). (d) Combining domain-elongation RDCs and MD simulations in the construction of atomic-resolution dynamic ensembles of TAR reveals conformations (in gray) very similar to those observed in ligand bound states (in orange) (Adapted from ref. by permission of Oxford University Press).
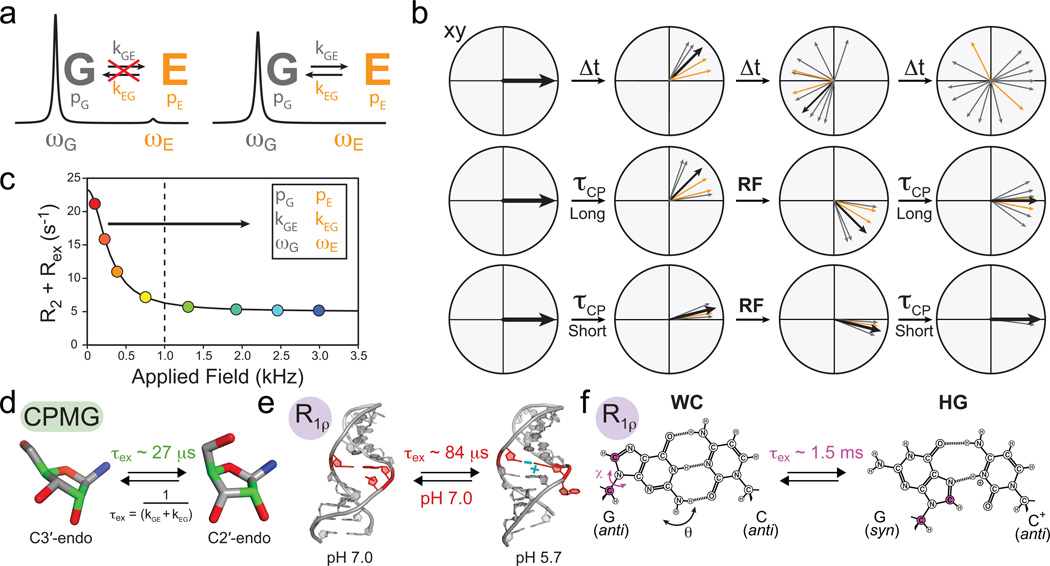
Characterizing micro- to millisecond exchange by relaxation dispersion. (a) Exchange between ground (G) and excited (E) states leads to broadening of the G signal and disappearance of the E signal. (b) Fanning out of bulk magnetization due to exchange can be suppressed by application of RF fields. (c) Characteristic relaxation dispersion curve showing the power dependence of Rex which can be used to extract in favorable cases the populations (p), exchange rates (k) and chemical shifts (ω) of G and E. (d–f) Examples of conformational exchange characterized by CPMG and R1ρ carbon relaxation dispersion. (d) Sugar repuckering in a GCAA tetraloop using selectively labeled C2' and C4' (green) probes. (e) Transition toward an excited state structure in the U6 RNA involving a CA+ base-pair and looping out of a uridine bulge. (f) Transition to Hoogsteen (HG) GC+ base-pairs in canonical duplex DNA (Adapted from ref. 70).
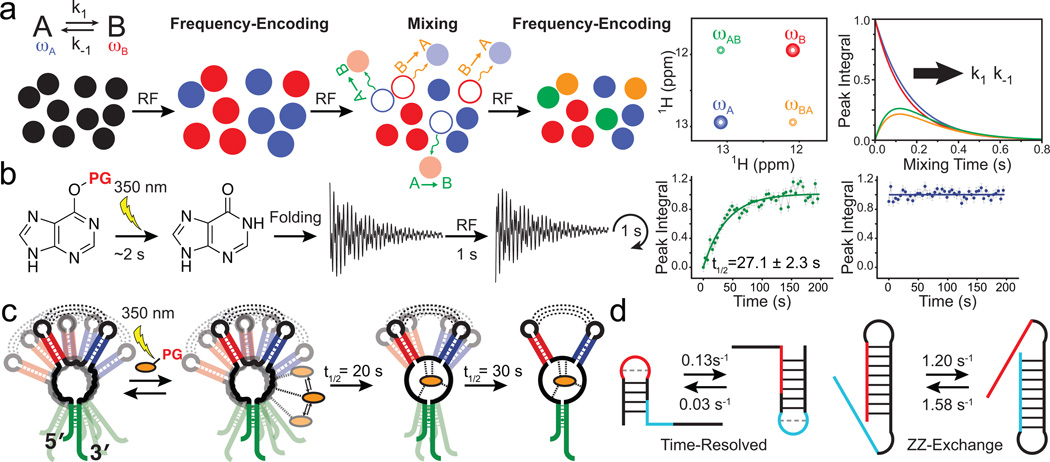
Characterizing conformational transitions occurring at millisecond and longer timescales by ZZ-exchange and time-resolved NMR. Schematic of the (a) ZZ-exchange and (b) time-resolved NMR experiment. (c) Visualizing a multi-step ligand-induced conformational transition in the guanine sensing riboswitch using laser induced time-resolved NMR (Adapted from ref. , copyright 2007 National Academy of Sciences, USA). (d) Characterizing slow and fast RNA refolding rates by time-resolved and ZZ-exchange NMR, respectively.
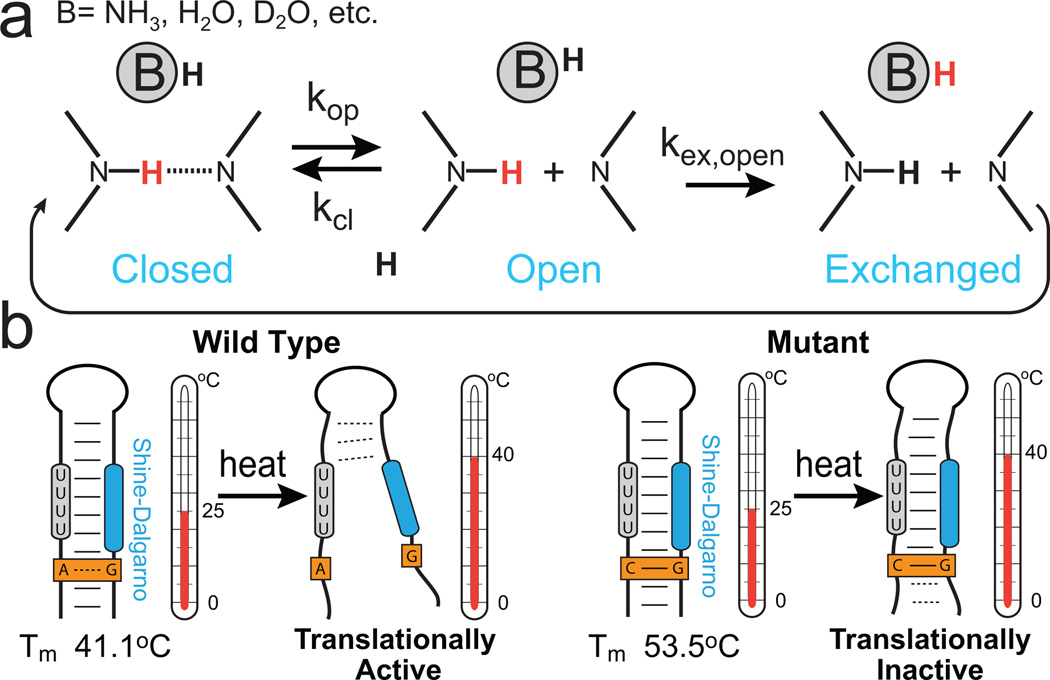
Characterizing base-pair opening dynamics by imino proton exchange. (a) Schematic of a two-step (direct or water-mediated) imino proton exchange for an RNA base pair catalyzed by a proton acceptor B, followed by reversible base-pair formation. H exchange experiments can be used to measure rate constants in each step. (b) Imino proton exchange study of the wild-type (left) Salmonella FourU RNA thermometer and its translationally inactive GA-to-GC mutant (right), showed a correlation between changes in local base-pair stabilities and changes in global melting profiles of the wild-type and mutant constructs.
References
-
- Frauenfelder H, Sligar SG, Wolynes PG. The Energy Landscapes and Motions of Proteins. Science. 1991;254:1598–1603. - PubMed
-
- Cruz JA, Westhof E. The Dynamic Landscapes of RNA Architecture. Cell. 2009;136:604–609. - PubMed
-
- Blount KF, Breaker RR. Riboswitches as antibacterial drug targets. Nat. Biotechnol. 2006;24:1558–1564. - PubMed
-
- Micura R, Hobartner C. On secondary structure rearrangements and equilibria of small RNAs. Chembiochem. 2003;4:984–990. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
