

Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals
- PMID: 22037601
- PMCID: PMC3242449
- DOI: 10.1038/ni.2143
Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals
Abstract
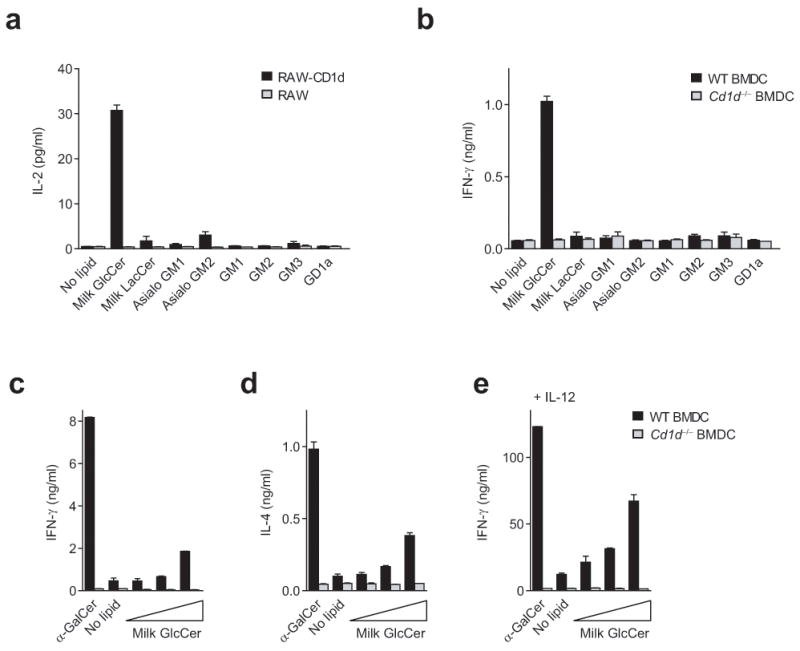
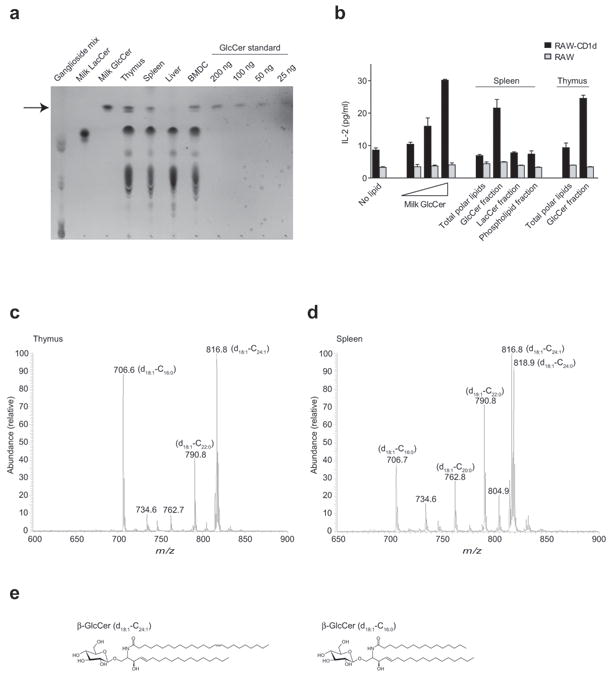
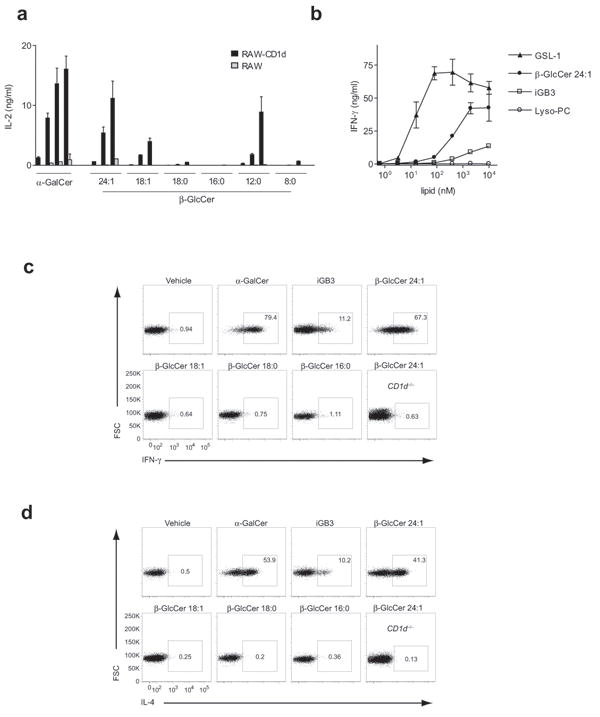
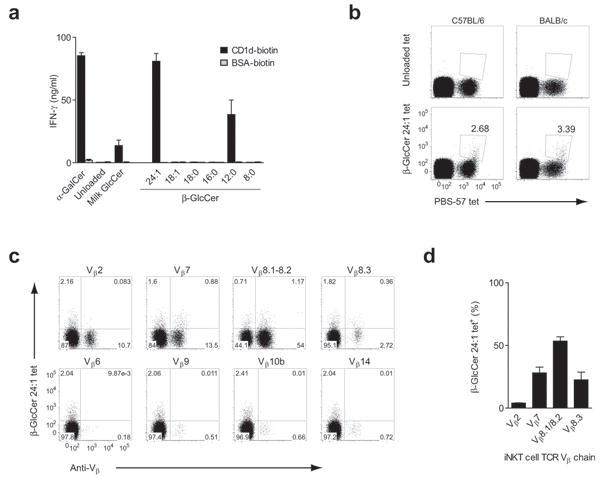
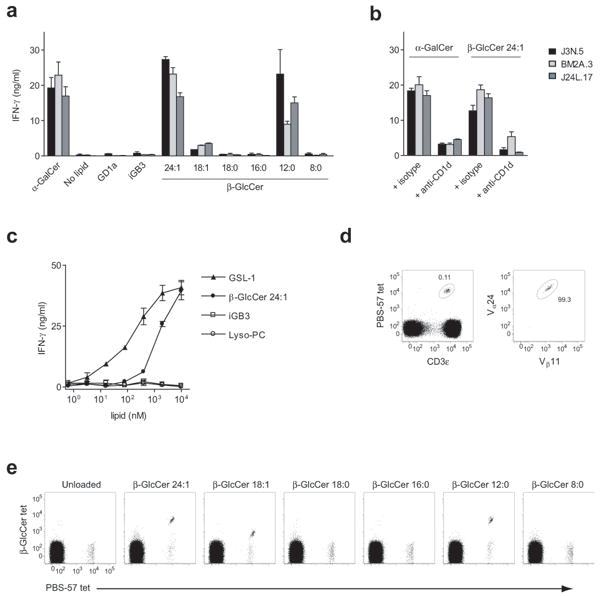
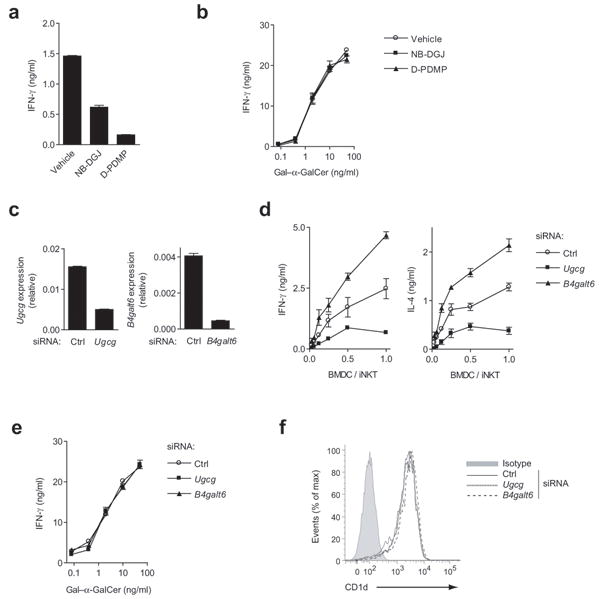
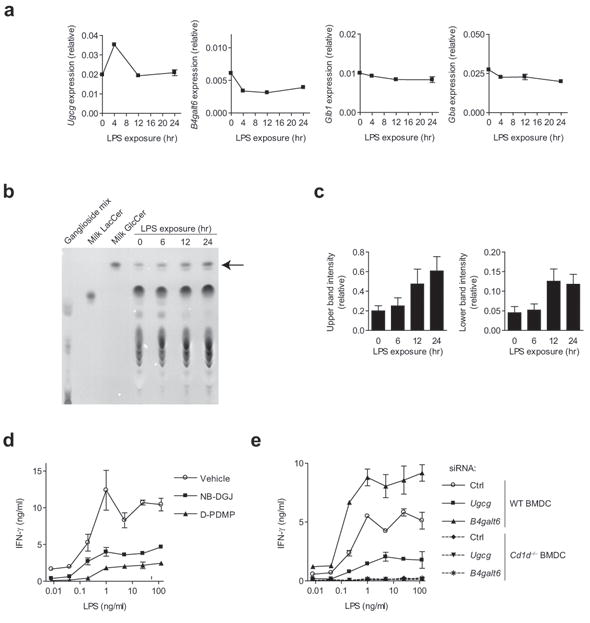
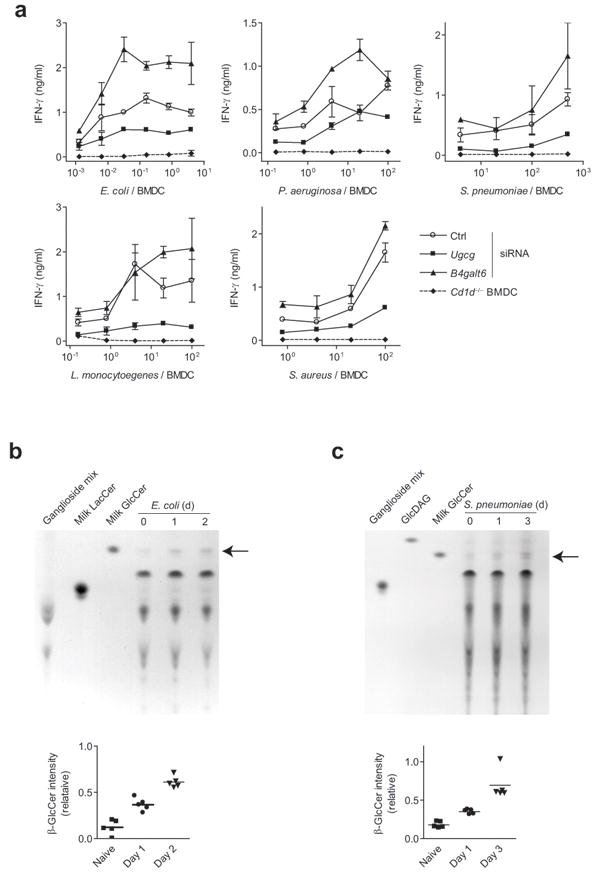
Invariant natural killer T cells (iNKT cells) have a prominent role during infection and other inflammatory processes, and these cells can be activated through their T cell antigen receptors by microbial lipid antigens. However, increasing evidence shows that they are also activated in situations in which foreign lipid antigens would not be present, which suggests a role for lipid self antigen. We found that an abundant endogenous lipid, β-D-glucopyranosylceramide (β-GlcCer), was a potent iNKT cell self antigen in mouse and human and that its activity depended on the composition of the N-acyl chain. Furthermore, β-GlcCer accumulated during infection and in response to Toll-like receptor agonists, contributing to iNKT cell activation. Thus, we propose that recognition of β-GlcCer by the invariant T cell antigen receptor translates innate danger signals into iNKT cell activation.
Figures
Comment in
-
Beta-testing NKT cell self-reactivity.Nat Immunol. 2011 Nov 16;12(12):1135-7. doi: 10.1038/ni.2162. Nat Immunol. 2011. PMID: 22089211 No abstract available.
References
-
- Brigl M, Brenner MB. CD1: antigen presentation and T cell function. Annu Rev Immunol. 2004;22:817–890. - PubMed
-
- Kawano T, et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science. 1997;278:1626–1629. - PubMed
-
- Kinjo Y, et al. Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat Immunol. 2006;7:978–986. - PubMed
-
- Kinjo Y, et al. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature. 2005;434:520–525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01AI063428/AI/NIAID NIH HHS/United States
- T32HL007633/HL/NHLBI NIH HHS/United States
- T32 HL007633/HL/NHLBI NIH HHS/United States
- K08 AI077795/AI/NIAID NIH HHS/United States
- P60-DK20579/DK/NIDDK NIH HHS/United States
- T32 AI007306/AI/NIAID NIH HHS/United States
- R01 AI063428/AI/NIAID NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- P41 RR005351/RR/NCRR NIH HHS/United States
- R01 AI028973/AI/NIAID NIH HHS/United States
- P41 RR000954/RR/NCRR NIH HHS/United States
- P41-RR05351/RR/NCRR NIH HHS/United States
- T32AI007306/AI/NIAID NIH HHS/United States
- P60 DK020579/DK/NIDDK NIH HHS/United States
- 084923/B/08/Z/WT_/Wellcome Trust/United Kingdom
- K08AI077795/AI/NIAID NIH HHS/United States
- 18892/ARC_/Arthritis Research UK/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- G0400421/MRC_/Medical Research Council/United Kingdom
- P41-RR00954/RR/NCRR NIH HHS/United States
- P30 AI060354/AI/NIAID NIH HHS/United States
- P30-DK56341/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
