

A novel mutation within the MIR96 gene causes non-syndromic inherited hearing loss in an Italian family by altering pre-miRNA processing
- PMID: 22038834
- PMCID: PMC3259013
- DOI: 10.1093/hmg/ddr493
A novel mutation within the MIR96 gene causes non-syndromic inherited hearing loss in an Italian family by altering pre-miRNA processing
Abstract
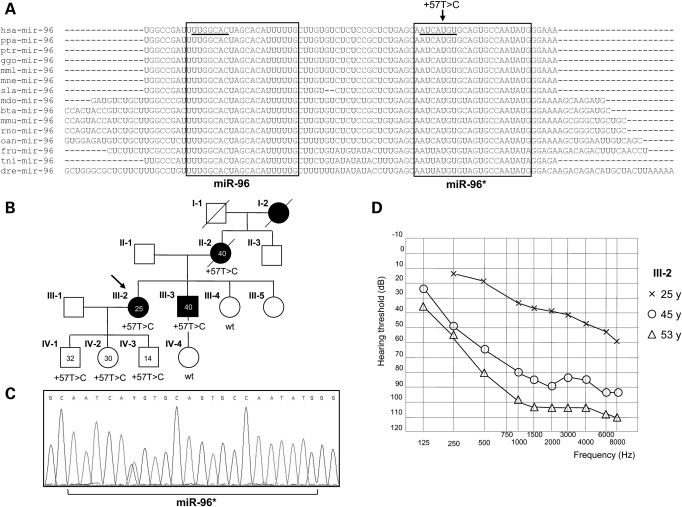
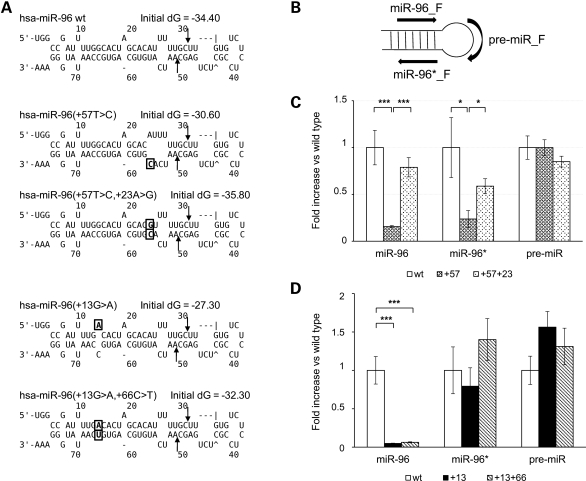
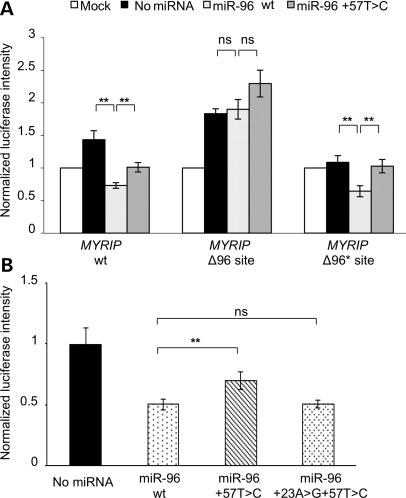
The miR-96, miR-182 and miR-183 microRNA (miRNA) family is essential for differentiation and function of the vertebrate inner ear. Recently, point mutations within the seed region of miR-96 were reported in two Spanish families with autosomal dominant non-syndromic sensorineural hearing loss (NSHL) and in a mouse model of NSHL. We screened 882 NSHL patients and 836 normal-hearing Italian controls and identified one putative novel mutation within the miR-96 gene in a family with autosomal dominant NSHL. Although located outside the mature miR-96 sequence, the detected variant replaces a highly conserved nucleotide within the companion miR-96*, and is predicted to reduce the stability of the pre-miRNA hairpin. To evaluate the effect of the detected mutation on miR-96/mir-96* biogenesis, we investigated the maturation of miR-96 by transient expression in mammalian cells, followed by real-time reverse-transcription polymerase chain reaction (PCR). We found that both miR-96 and miR-96* levels were significantly reduced in the mutant, whereas the precursor levels were unaffected. Moreover, miR-96 and miR-96* expression levels could be restored by a compensatory mutation that reconstitutes the secondary structure of the pre-miR-96 hairpin, demonstrating that the mutation hinders precursor processing, probably interfering with Dicer cleavage. Finally, even though the mature miR-96 sequence is not altered, we demonstrated that the identified mutation significantly impacts on miR-96 regulation of selected targets. In conclusion, we provide further evidence of the involvement of miR-96 mutations in human deafness and demonstrate that a quantitative defect of this miRNA may contribute to NSHL.
Figures
References
-
- Huntzinger E., Izaurralde E. Gene silencing by microRNAs: contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011;12:99–110. doi:10.1038/nrg2936. - DOI - PubMed
-
- Bartel D.P. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–233. doi:10.1016/j.cell.2009.01.002. - DOI - PMC - PubMed
-
- Krol J., Loedige I., Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010;11:597–610. - PubMed
-
- Ro S., Park C., Young D., Sanders K.M., Yan W. Tissue-dependent paired expression of miRNAs. Nucleic Acids Res. 2007;35:5944–5953. doi:10.1093/nar/gkm641. - DOI - PMC - PubMed
-
- Okamura K., Phillips M., Tyler D., Duan H., Chou Y., Lai E. The regulatory activity of microRNA* species has substantial influence on microRNA and 3′ UTR evolution. Nat. Struct. Mol. Biol. 2008;15:354–363. doi:10.1038/nsmb.1409. - DOI - PMC - PubMed
