

Characterization of Brca2-deficient plants excludes the role of NHEJ and SSA in the meiotic chromosomal defect phenotype
- PMID: 22039535
- PMCID: PMC3198793
- DOI: 10.1371/journal.pone.0026696
Characterization of Brca2-deficient plants excludes the role of NHEJ and SSA in the meiotic chromosomal defect phenotype
Abstract
In somatic cells, three major pathways are involved in the repair of DNA double-strand breaks (DBS): Non-Homologous End Joining (NHEJ), Single-Strand Annealing (SSA) and Homologous Recombination (HR). In somatic and meiotic HR, DNA DSB are 5' to 3' resected, producing long 3' single-stranded DNA extensions. Brca2 is essential to load the Rad51 recombinase onto these 3' overhangs. The resulting nucleofilament can thus invade a homologous DNA sequence to copy and restore the original genetic information. In Arabidopsis, the inactivation of Brca2 specifically during meiosis by an RNAi approach results in aberrant chromosome aggregates, chromosomal fragmentation and missegregation leading to a sterility phenotype. We had previously suggested that such chromosomal behaviour could be due to NHEJ. In this study, we show that knock-out plants affected in both BRCA2 genes show the same meiotic phenotype as the RNAi-inactivated plants. Moreover, it is demonstrated that during meiosis, neither NHEJ nor SSA compensate for HR deficiency in BRCA2-inactivated plants. The role of the plant-specific DNA Ligase6 is also excluded. The possible mechanism(s) involved in the formation of these aberrant chromosomal bridges in the absence of HR during meiosis are discussed.
Conflict of interest statement
Figures
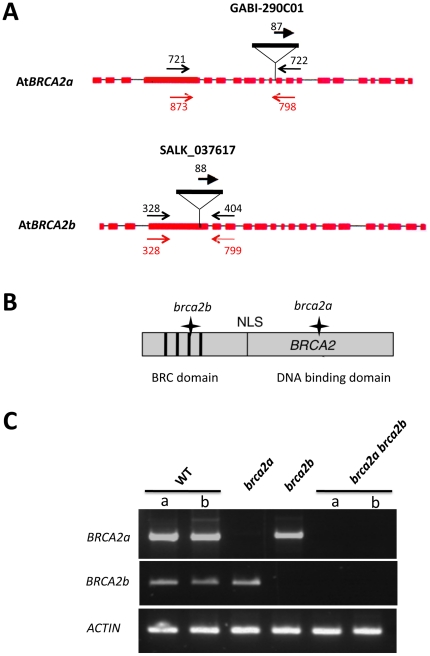
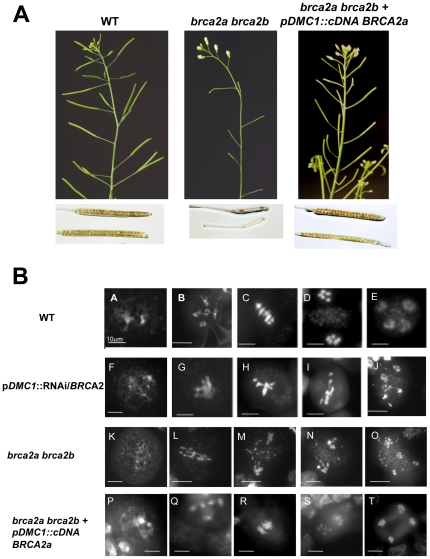
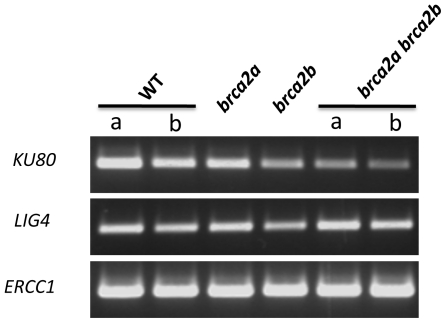
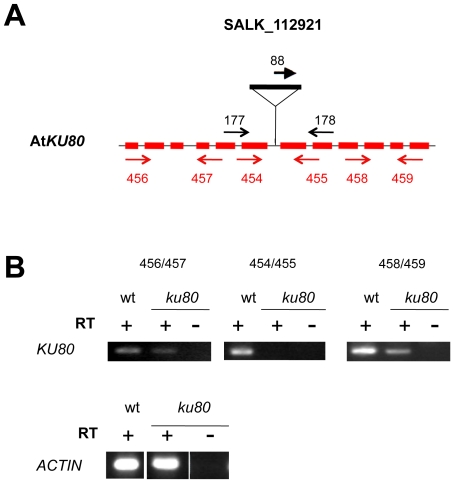
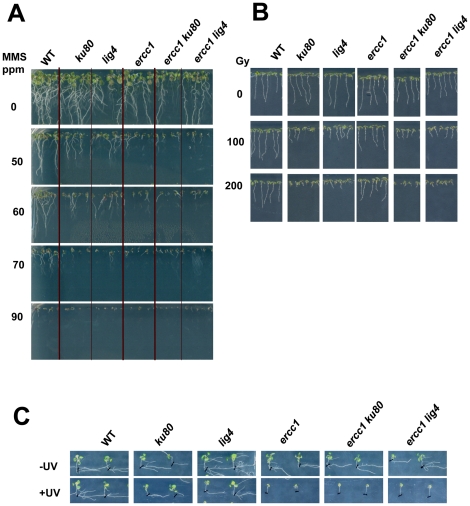
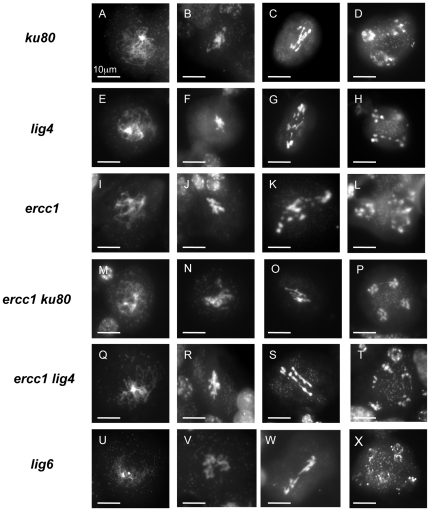
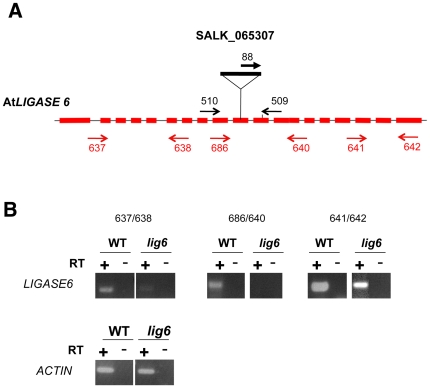
References
-
- Daley JM, Palmbos PL, Wu D, Wilson TE. Nonhomologous end joining in yeast. Annu Rev Genet. 2005;39:431–451. - PubMed
-
- Tamura K, Adachi Y, Chiba K, Oguchi K, Takahashi H. Identification of Ku70 and Ku80 homologues in Arabidopsis thaliana: evidence for a role in the repair of DNA double-strand breaks. Plant J. 2002;29:771–781. - PubMed
-
- Friesner J, Britt AB. Ku80- and DNA ligase IV-deficient plants are sensitive to ionizing radiation and defective in T-DNA integration. Plant J. 2003;34:427–440. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
