

CcpA ensures optimal metabolic fitness of Streptococcus pneumoniae
- PMID: 22039538
- PMCID: PMC3198803
- DOI: 10.1371/journal.pone.0026707
CcpA ensures optimal metabolic fitness of Streptococcus pneumoniae
Abstract
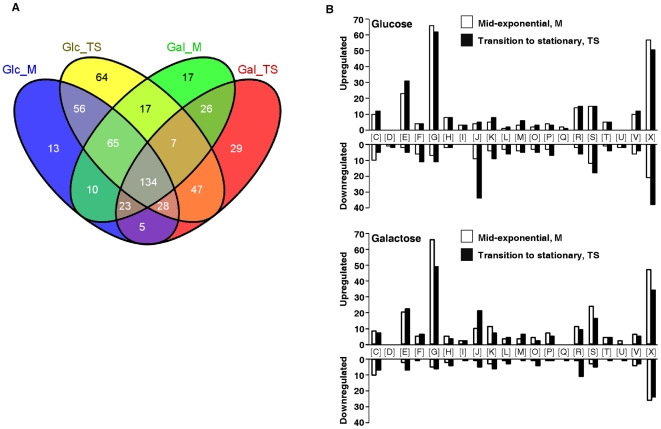
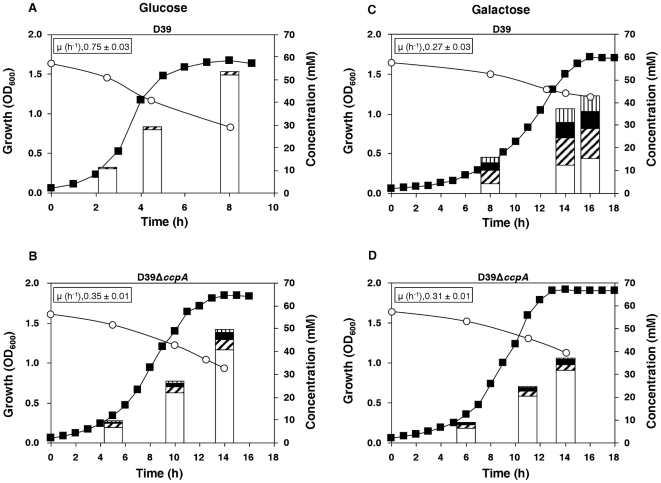
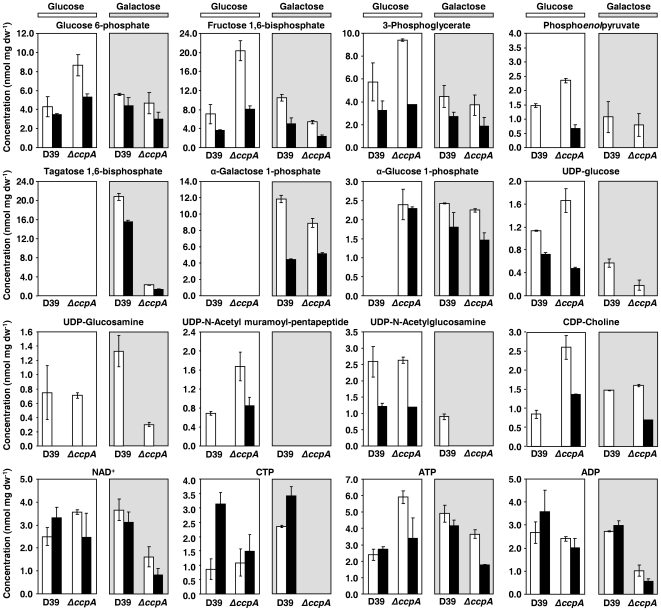
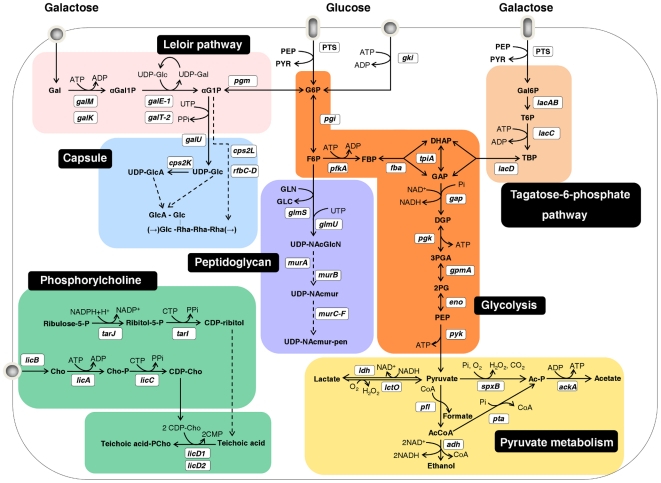
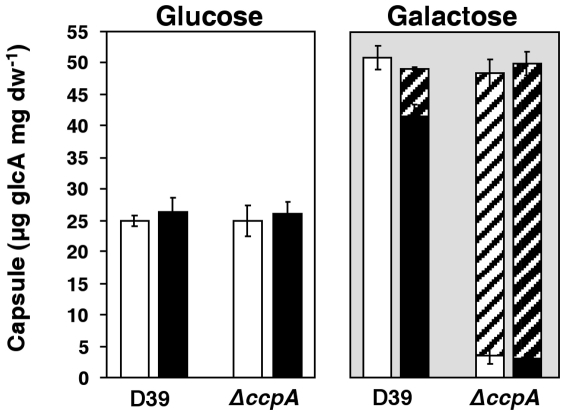
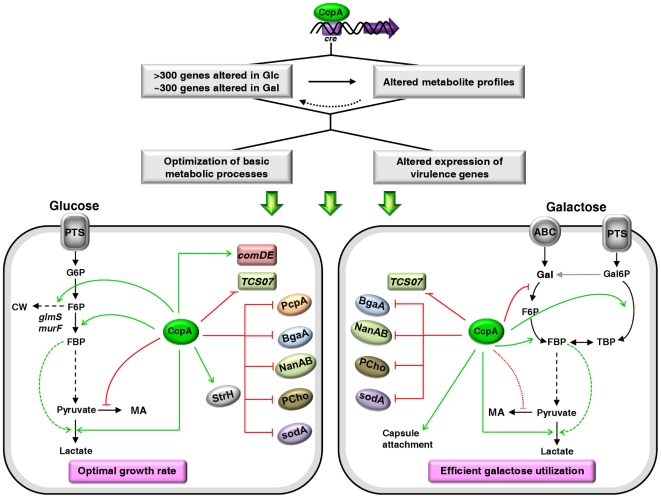
In gram-positive bacteria, the transcriptional regulator CcpA is at the core of catabolite control mechanisms. In the human pathogen Streptococcus pneumoniae, links between CcpA and virulence have been established, but its role as a master regulator in different nutritional environments remains to be elucidated. Thus, we performed whole-transcriptome and metabolic analyses of S. pneumoniae D39 and its isogenic ccpA mutant during growth on glucose or galactose, rapidly and slowly metabolized carbohydrates presumably encountered by the bacterium in different host niches. CcpA affected the expression of up to 19% of the genome covering multiple cellular processes, including virulence, regulatory networks and central metabolism. Its prevalent function as a repressor was observed on glucose, but unexpectedly also on galactose. Carbohydrate-dependent CcpA regulation was also observed, as for the tagatose 6-phosphate pathway genes, which were activated by galactose and repressed by glucose. Metabolite analyses revealed that two pathways for galactose catabolism are functionally active, despite repression of the Leloir genes by CcpA. Surprisingly, galactose-induced mixed-acid fermentation apparently required CcpA, since genes involved in this type of metabolism were mostly under CcpA-repression. These findings indicate that the role of CcpA extends beyond transcriptional regulation, which seemingly is overlaid by other regulatory mechanisms. In agreement, CcpA influenced the level of many intracellular metabolites potentially involved in metabolic regulation. Our data strengthen the view that a true understanding of cell physiology demands thorough analyses at different cellular levels. Moreover, integration of transcriptional and metabolic data uncovered a link between CcpA and the association of surface molecules (e.g. capsule) to the cell wall. Hence, CcpA may play a key role in mediating the interaction of S. pneumoniae with its host. Overall, our results support the hypothesis that S. pneumoniae optimizes basic metabolic processes, likely enhancing in vivo fitness, in a CcpA-mediated manner.
Conflict of interest statement
Figures

References
-
- Mitchell TJ. The pathogenesis of streptococcal infections: from tooth decay to meningitis. Nat Rev Microbiol. 2003;1:219–230. - PubMed
-
- Kadioglu A, Weiser JN, Paton JC, Andrew PW. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat Rev Microbiol. 2008;6:288–301. - PubMed
-
- Görke B, Stülke J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat Rev Microbiol. 2008;6:613–624. - PubMed
-
- Lulko AT, Buist G, Kok J, Kuipers OP. Transcriptome analysis of temporal regulation of carbon metabolism by CcpA in Bacillus subtilis reveals additional target genes. J Mol Microbiol Biotechnol. 2007;12:82–95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
