

FGF2 gene transfer restores hippocampal functions in mouse models of Alzheimer's disease and has therapeutic implications for neurocognitive disorders
- PMID: 22042871
- PMCID: PMC3241747
- DOI: 10.1073/pnas.1102349108
FGF2 gene transfer restores hippocampal functions in mouse models of Alzheimer's disease and has therapeutic implications for neurocognitive disorders
Erratum in
- Proc Natl Acad Sci U S A. 2011 Dec 27;108(52):21282
Abstract
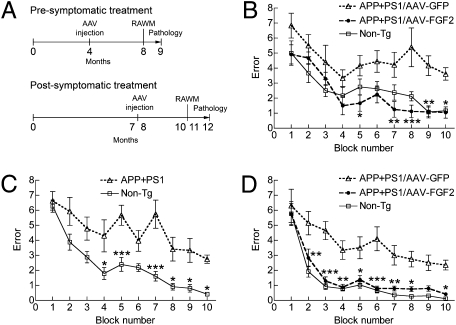
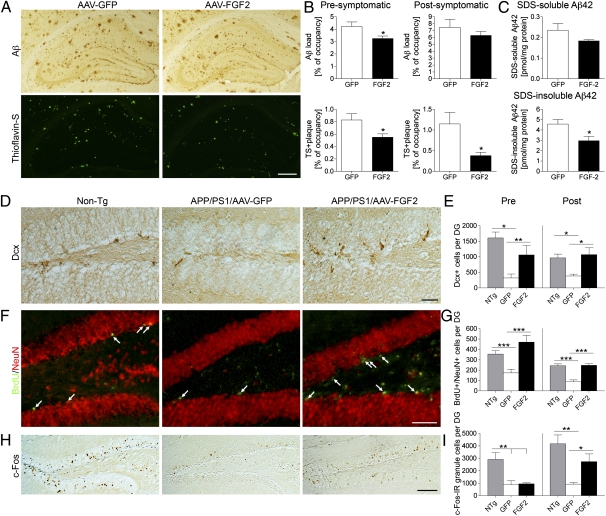
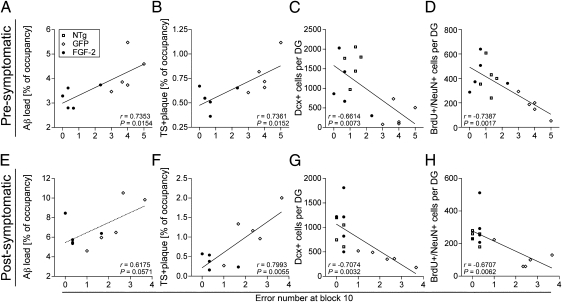
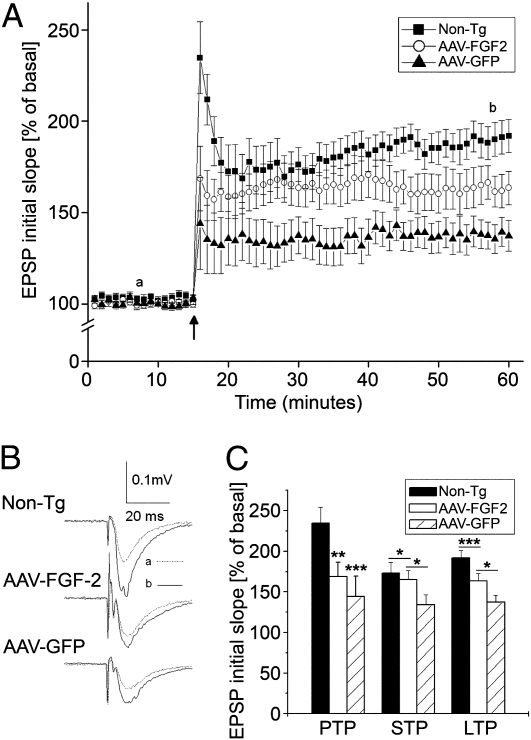
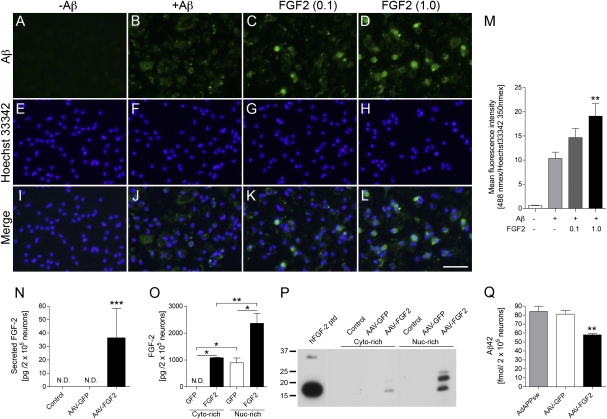
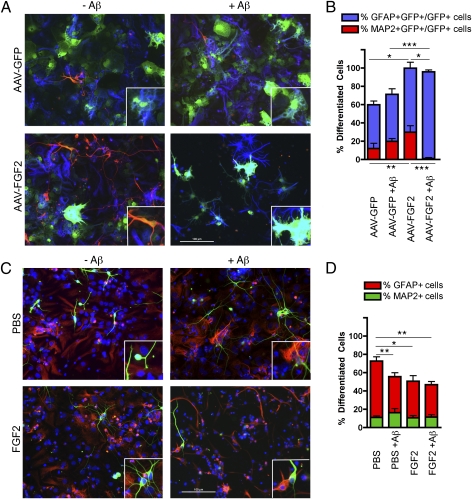
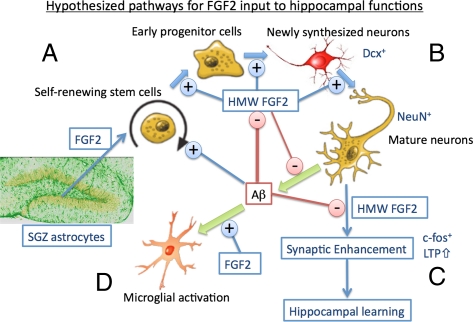
The adult hippocampus plays a central role in memory formation, synaptic plasticity, and neurogenesis. The subgranular zone of the dentate gyrus contains neural progenitor cells with self-renewal and multilineage potency. Transgene expression of familial Alzheimer's disease-linked mutants of β-amyloid precursor protein (APP) and presenilin-1 leads to a significant inhibition of neurogenesis, which is potentially linked to age-dependent memory loss. To investigate the effect of neurogenesis on cognitive function in a relevant disease model, FGF2 gene is delivered bilaterally to the hippocampi of APP+presenilin-1 bigenic mice via an adenoassociated virus serotype 2/1 hybrid (AAV2/1-FGF2). Animals injected with AAV2/1-FGF2 at a pre- or postsymptomatic stage show significantly improved spatial learning in the radial arm water maze test. A neuropathological investigation demonstrates that AAV2/1-FGF2 injection enhances the number of doublecortin, BrdU/NeuN, and c-fos-positive cells in the dentate gyrus, and the clearance of fibrillar amyloid-β peptide (Aβ) in the hippocampus. AAV2/1-FGF2 injection also enhances long-term potentiation in another APP mouse model (J20) compared with control AAV2/1-GFP-injected littermates. An in vitro study confirmed the enhanced neurogenesis of mouse neural stem cells by direct AAV2/1-FGF2 infection in an Aβ oligomer-sensitive manner. Further, FGF2 enhances Aβ phagocytosis in primary cultured microglia, and reduces Aβ production from primary cultured neurons after AAV2/1-FGF2 infection. Thus, our data indicate that virus-mediated FGF2 gene delivery has potential as an alternative therapy of Alzheimer's disease and possibly other neurocognitive disorders.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Martin JB. Molecular basis of the neurodegenerative disorders. N Engl J Med. 1999;340:1970–1980. - PubMed
-
- Forman MS, Trojanowski JQ, Lee VM. Neurodegenerative diseases: A decade of discoveries paves the way for therapeutic breakthroughs. Nat Med. 2004;10:1055–1063. - PubMed
-
- Selkoe DJ. Alzheimer's disease is a synaptic failure. Science. 2002;298:789–791. - PubMed
-
- Blennow K, de Leon MJ, Zetterberg H. Alzheimer's disease. Lancet. 2006;368:387–403. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
