

Upregulation of micro RNA-146a (miRNA-146a), a marker for inflammatory neurodegeneration, in sporadic Creutzfeldt-Jakob disease (sCJD) and Gerstmann-Straussler-Scheinker (GSS) syndrome
- PMID: 22043907
- PMCID: PMC3719866
- DOI: 10.1080/15287394.2011.618973
Upregulation of micro RNA-146a (miRNA-146a), a marker for inflammatory neurodegeneration, in sporadic Creutzfeldt-Jakob disease (sCJD) and Gerstmann-Straussler-Scheinker (GSS) syndrome
Abstract
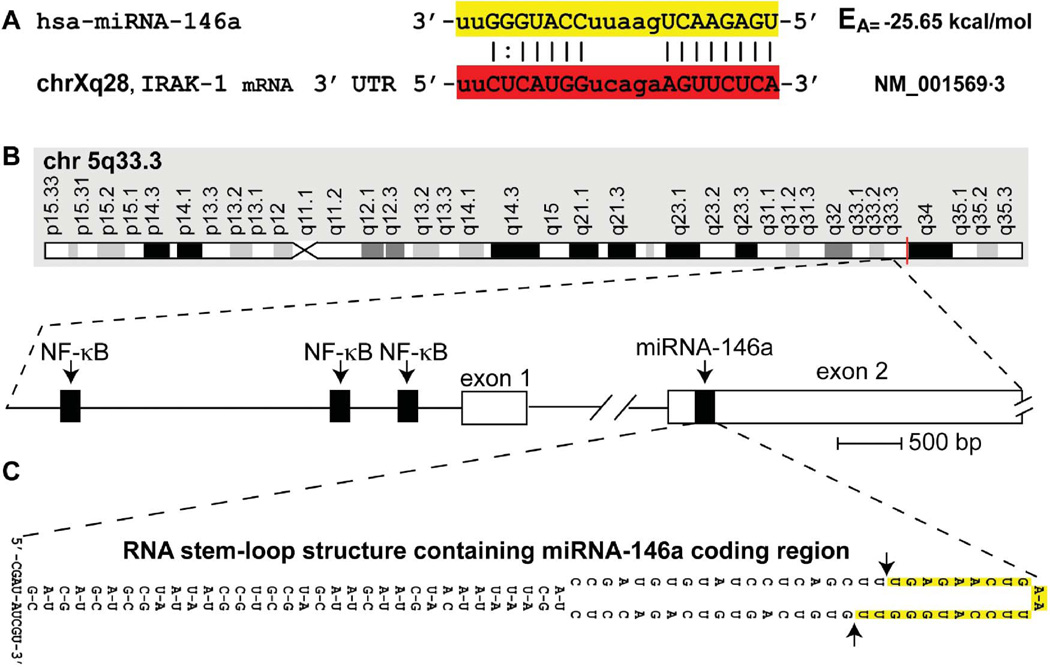
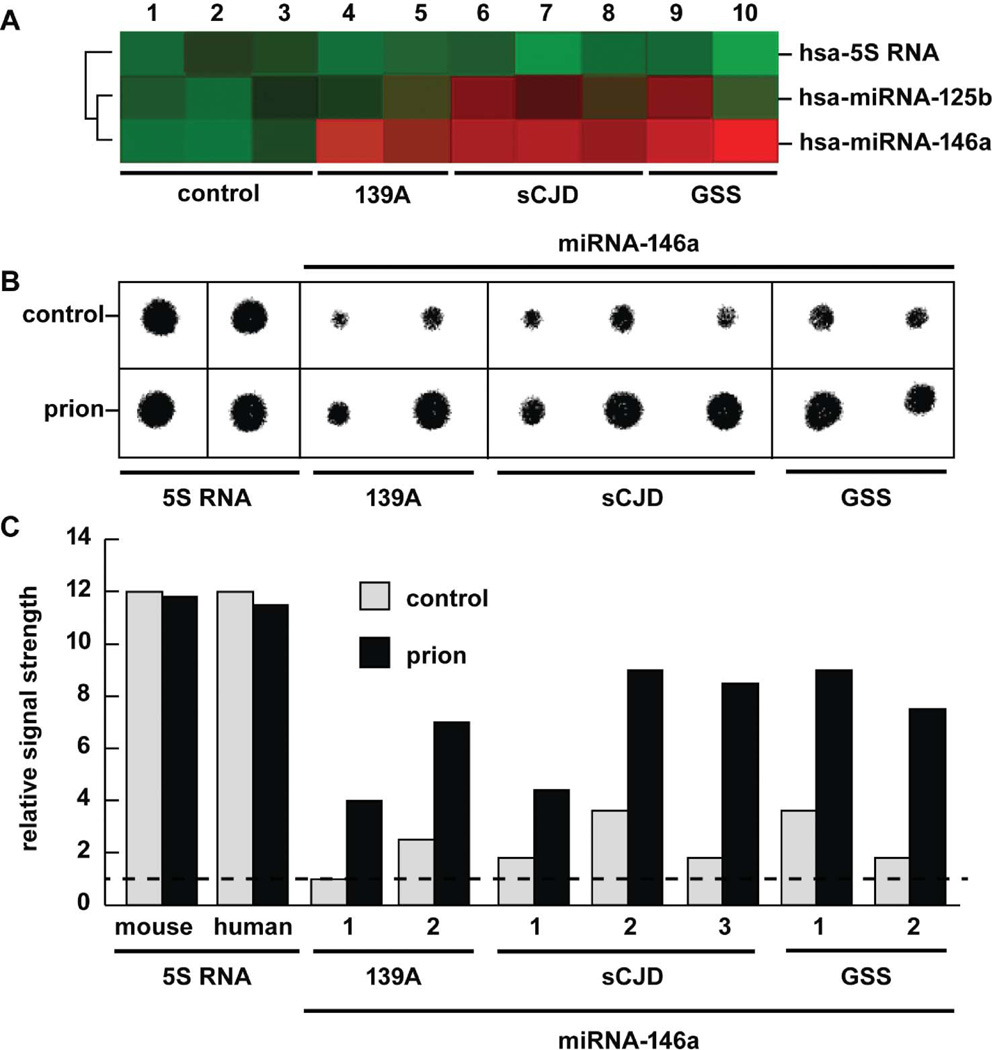
A mouse- and human-brain-abundant, nuclear factor (NF)-кB-regulated, micro RNA-146a (miRNA-146a) is an important modulator of the innate immune response and inflammatory signaling in specific immunological and brain cell types. Levels of miRNA-146a are induced in human brain cells challenged with at least five different species of single- or double-stranded DNA or RNA neurotrophic viruses, suggesting a broad role for miRNA-146a in the brain's innate immune response and antiviral immunity. Upregulated miRNA-146a is also observed in pro-inflammatory cytokine-, Aβ42 peptide- and neurotoxic metal-induced, oxidatively stressed human neuronal-glial primary cell cocultures, in murine scrapie and in Alzheimer's disease (AD) brain. In AD, miRNA-146a levels are found to progressively increase with disease severity and co-localize to brain regions enriched in inflammatory neuropathology. This study provides evidence of upregulation of miRNA-146a in extremely rare (incidence 1-10 per 100 million) human prion-based neurodegenerative disorders, including sporadic Creutzfeldt-Jakob disease (sCJD) and Gerstmann-Straussler-Scheinker syndrome (GSS). The findings suggest that an upregulated miRNA-146a may be integral to innate immune or inflammatory brain cell responses in prion-mediated infections and to progressive and irreversible neurodegeneration of both the murine and human brain.
Figures
Similar articles
-
microRNA-146a-5p, Neurotropic Viral Infection and Prion Disease (PrD).Int J Mol Sci. 2021 Aug 25;22(17):9198. doi: 10.3390/ijms22179198. Int J Mol Sci. 2021. PMID: 34502105 Free PMC article. Review.
-
Ultrastructural study of florid plaques in variant Creutzfeldt-Jakob disease: a comparison with amyloid plaques in kuru, sporadic Creutzfeldt-Jakob disease and Gerstmann-Sträussler-Scheinker disease.Neuropathol Appl Neurobiol. 2009 Feb;35(1):46-59. doi: 10.1111/j.1365-2990.2008.00959.x. Epub 2008 May 30. Neuropathol Appl Neurobiol. 2009. PMID: 18513219
-
Gerstmann-Sträussler-Scheinker disease and "anchorless prion protein" mice share prion conformational properties diverging from sporadic Creutzfeldt-Jakob disease.J Biol Chem. 2014 Feb 21;289(8):4870-81. doi: 10.1074/jbc.M113.531335. Epub 2014 Jan 7. J Biol Chem. 2014. PMID: 24398683 Free PMC article.
-
microRNA-146a as a biomarker for transmissible spongiform encephalopathy.Folia Neuropathol. 2022;60(1):24-34. doi: 10.5114/fn.2022.113561. Folia Neuropathol. 2022. PMID: 35359143
-
[Creutzfeldt-Jakob disease(CJD) and Gerstmann-Sträussler-Scheinker syndrome(GSS)].Nihon Rinsho. 1997 Apr;55(4):972-7. Nihon Rinsho. 1997. PMID: 9103904 Review. Japanese.
Cited by
-
Identification of circulating microRNA signatures as potential biomarkers in the serum of elk infected with chronic wasting disease.Sci Rep. 2019 Dec 23;9(1):19705. doi: 10.1038/s41598-019-56249-6. Sci Rep. 2019. PMID: 31873177 Free PMC article.
-
Panoramic Visualization of Circulating MicroRNAs Across Neurodegenerative Diseases in Humans.Mol Neurobiol. 2019 Nov;56(11):7380-7407. doi: 10.1007/s12035-019-1615-1. Epub 2019 Apr 29. Mol Neurobiol. 2019. PMID: 31037649 Free PMC article.
-
Identification of endogenous reference genes for the analysis of microRNA expression in the hippocampus of the pilocarpine-induced model of mesial temporal lobe epilepsy.PLoS One. 2014 Jun 25;9(6):e100529. doi: 10.1371/journal.pone.0100529. eCollection 2014. PLoS One. 2014. PMID: 24964029 Free PMC article.
-
MicroRNAs in Prion Diseases-From Molecular Mechanisms to Insights in Translational Medicine.Cells. 2021 Jun 29;10(7):1620. doi: 10.3390/cells10071620. Cells. 2021. PMID: 34209482 Free PMC article. Review.
-
Extracellular Vesicles, Stem Cells and the Role of miRNAs in Neurodegeneration.Curr Neuropharmacol. 2022;20(8):1450-1478. doi: 10.2174/1570159X19666210817150141. Curr Neuropharmacol. 2022. PMID: 34414870 Free PMC article. Review.
References
-
- Alexandrov PN, Zhao Y, Pogue AI, Tarr MA, Kruck TP, Percy ME, Cui JG, Lukiw WJ. Synergistic effects of iron and aluminum on stress-related gene expression in primary human neural cells. J. Alzheimers Dis. 2005;8:117–127. - PubMed
-
- Aronica E, Fluiter K, Iyer A, Zurolo E, Vreijling J, van Vliet EA, Baayen JC, Gorter JA. Expression pattern of miR-146a, an inflammation-associated microRNA, in experimental and human temporal lobe epilepsy. Eur. J. Neurosci. 2010;31:1100–1107. - PubMed
-
- Burmistrova OA, Goltsov AY, Abramova LI, Kaleda VG, Orlova VA, Rogaev EI. MicroRNA in schizophrenia: Genetic and expression analysis of miR-130b (22q11) Biochemistry (Moscow) 2007;72:578–582. - PubMed
-
- Collins S, McLean CA, Masters CL. Gerstmann–Sträussler–Scheinker syndrome, fatal familial insomnia, and kuru: A review of these less common human transmissible spongiform encephalopathies. J. Clin. Neurosci. 2001;8:387–397. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous