

Cholesterol esterification by ACAT2 is essential for efficient intestinal cholesterol absorption: evidence from thoracic lymph duct cannulation
- PMID: 22045928
- PMCID: PMC3243485
- DOI: 10.1194/jlr.M018820
Cholesterol esterification by ACAT2 is essential for efficient intestinal cholesterol absorption: evidence from thoracic lymph duct cannulation
Abstract
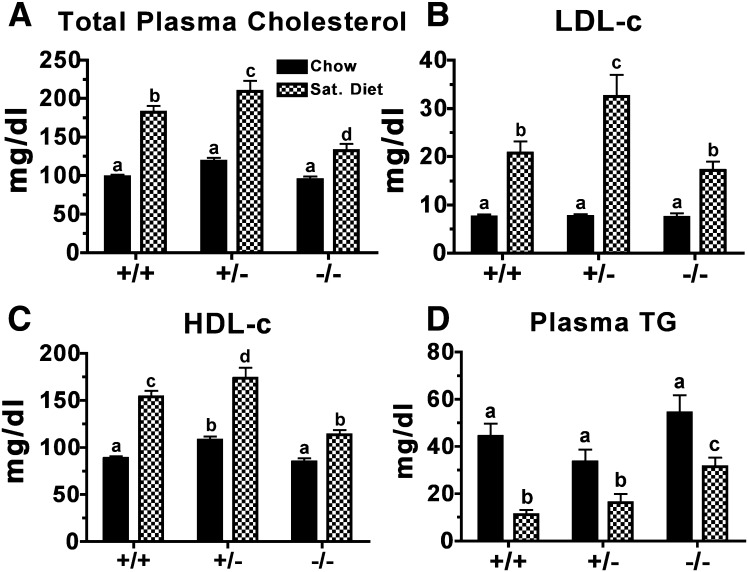
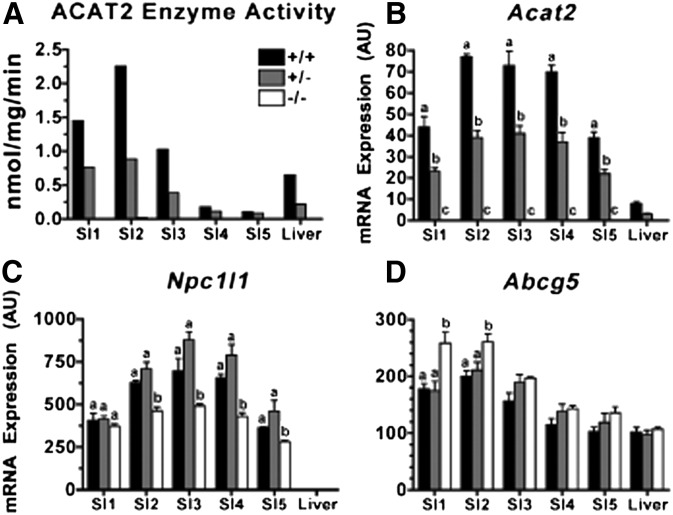
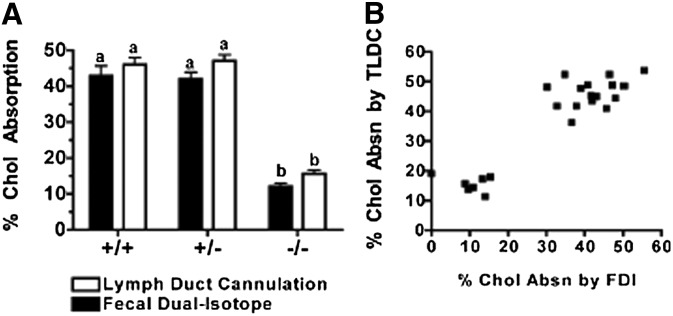
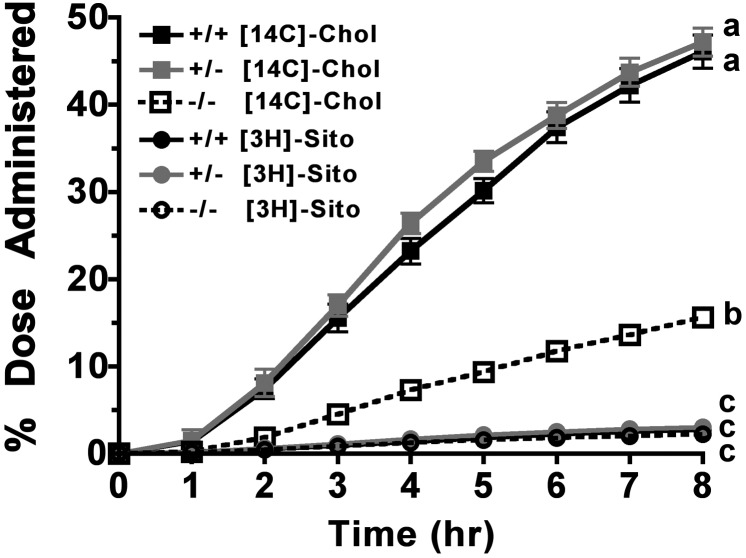
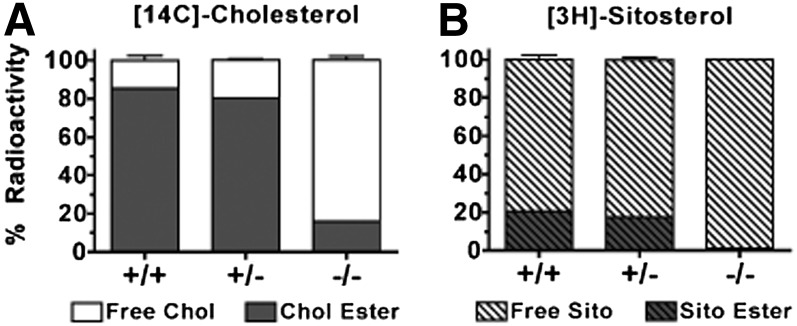
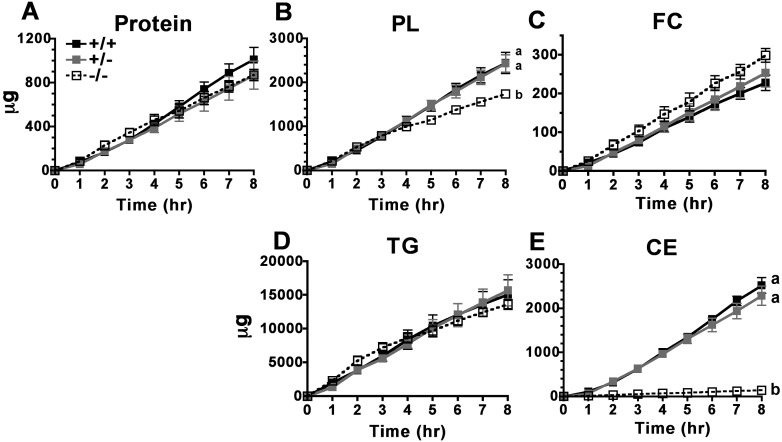
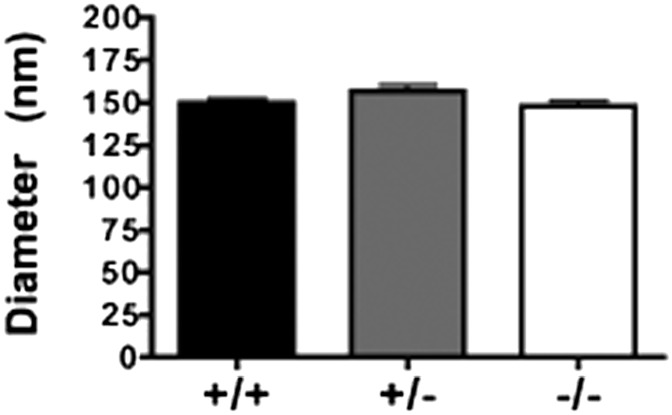
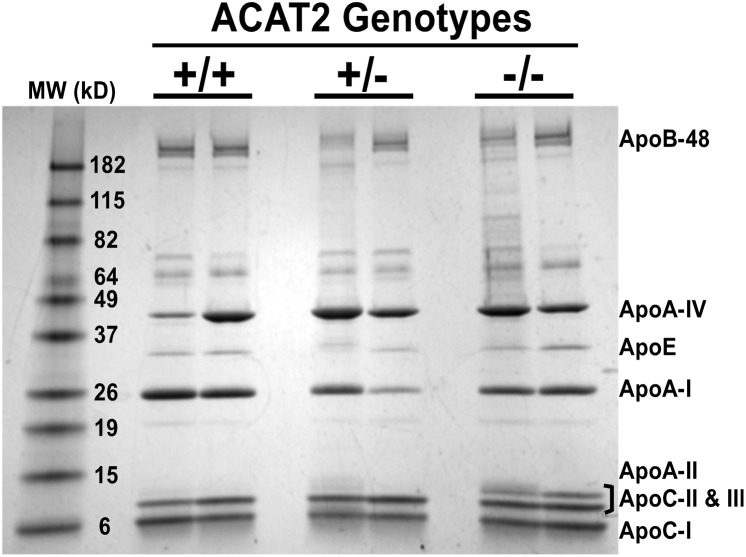
The hypothesis tested in this study was that cholesterol esterification by ACAT2 would increase cholesterol absorption efficiency by providing cholesteryl ester (CE) for incorporation into chylomicrons. The assumption was that absorption would be proportional to Acat2 gene dosage. Male ACAT2⁺/⁺, ACAT2⁺/⁻, and ACAT2⁻/⁻ mice were fed a diet containing 20% of energy as palm oil with 0.2% (w/w) cholesterol. Cholesterol absorption efficiency was measured by fecal dual-isotope and thoracic lymph duct cannulation (TLDC) methods using [³H]sitosterol and [¹⁴C]cholesterol tracers. Excellent agreement among individual mice was found for cholesterol absorption measured by both techniques. Cholesterol absorption efficiency in ACAT2⁻/⁻ mice was 16% compared with 46-47% in ACAT2⁺/⁺ and ACAT2⁺/⁻ mice. Chylomicrons from ACAT2⁺/⁺ and ACAT2⁺/⁻ mice carried ∼80% of total sterol mass as CE, whereas ACAT2⁻/⁻ chylomicrons carried >90% of sterol mass in the unesterified form. The total percentage of chylomicron mass as CE was reduced from 12% in the presence of ACAT2 to ∼1% in ACAT2⁻/⁻ mice. Altogether, the data demonstrate that ACAT2 increases cholesterol absorption efficiency by providing CE for chylomicron transport, but one copy of the Acat2 gene, providing ∼50% of ACAT2 mRNA and enzyme activity, was as effective as two copies in promoting cholesterol absorption.
Figures
References
-
- Rosamond W., Flegal K., Furie K., Go A., Greenlund K., Haase N., Hailpern S. M., Ho M., Howard V., Kissela B., et al. 2008. Heart disease and stroke statistics--2008 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation. 117: e25–e146. - PubMed
-
- Kannel W. B., Castelli W. P., Gordon T., McNamara P. M. 1971. Serum cholesterol, lipoproteins, and the risk of coronary heart disease. The Framingham study. Ann. Intern. Med. 74: 1–12. - PubMed
-
- Wang D. Q. 2007. Regulation of intestinal cholesterol absorption. Annu. Rev. Physiol. 69: 221–248. - PubMed
-
- McMurry M. P., Connor W. E., Lin D. S., Cerqueira M. T., Connor S. L. 1985. The absorption of cholesterol and the sterol balance in the Tarahumara Indians of Mexico fed cholesterol-free and high cholesterol diets. Am. J. Clin. Nutr. 41: 1289–1298. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
