

Biogenesis and functions of lipid droplets in plants: Thematic Review Series: Lipid Droplet Synthesis and Metabolism: from Yeast to Man
- PMID: 22045929
- PMCID: PMC3269164
- DOI: 10.1194/jlr.R021436
Biogenesis and functions of lipid droplets in plants: Thematic Review Series: Lipid Droplet Synthesis and Metabolism: from Yeast to Man
Abstract
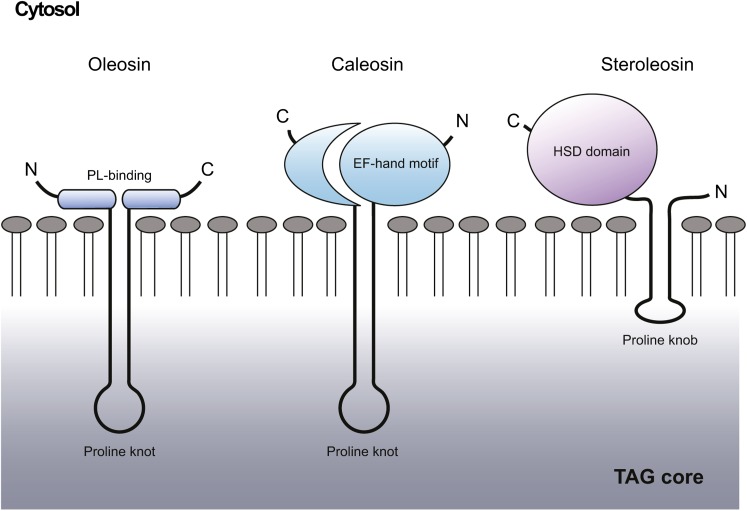
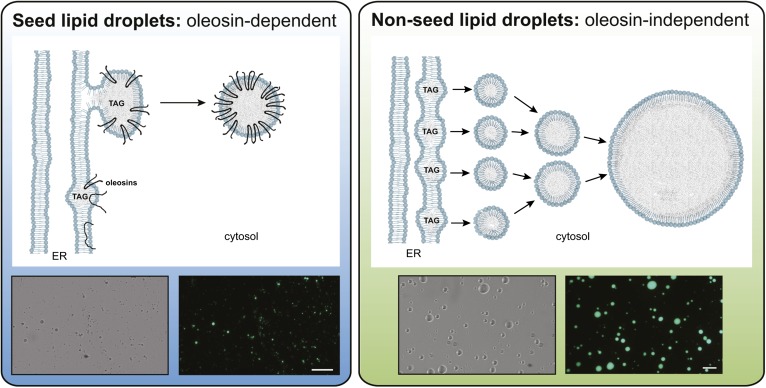
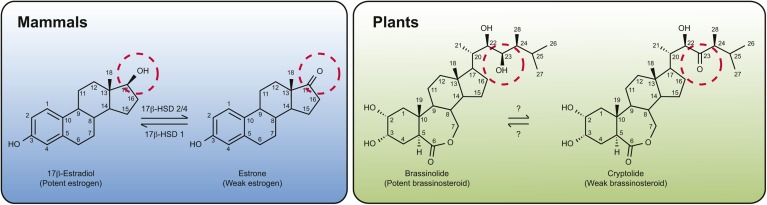
The compartmentation of neutral lipids in plants is mostly associated with seed tissues, where triacylglycerols (TAGs) stored within lipid droplets (LDs) serve as an essential physiological energy and carbon reserve during postgerminative growth. However, some nonseed tissues, such as leaves, flowers and fruits, also synthesize and store TAGs, yet relatively little is known about the formation or function of LDs in these tissues. Characterization of LD-associated proteins, such as oleosins, caleosins, and sterol dehydrogenases (steroleosins), has revealed surprising features of LD function in plants, including stress responses, hormone signaling pathways, and various aspects of plant growth and development. Although oleosin and caleosin proteins are specific to plants, LD-associated sterol dehydrogenases also are present in mammals, and in both plants and mammals these enzymes have been shown to be important in (steroid) hormone metabolism and signaling. In addition, several other proteins known to be important in LD biogenesis in yeasts and mammals are conserved in plants, suggesting that at least some aspects of LD biogenesis and/or function are evolutionarily conserved.
Figures
References
-
- Murphy D. J. The dynamic roles of intracellular lipid droplets: from archaea to mammals. Protoplasma. Epub ahead of print. October 15, 2011. - PubMed
-
- Frey-Wyssling A., Grieshaber E., Mühlethaler K. 1963. Origin of spherosomes in plant cells. J. Ultrastruct. Res. 8: 506–516.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
